

Identification of neutral tumor evolution across cancer types
- PMID: 26780609
- PMCID: PMC4934603
- DOI: 10.1038/ng.3489
Identification of neutral tumor evolution across cancer types
Abstract
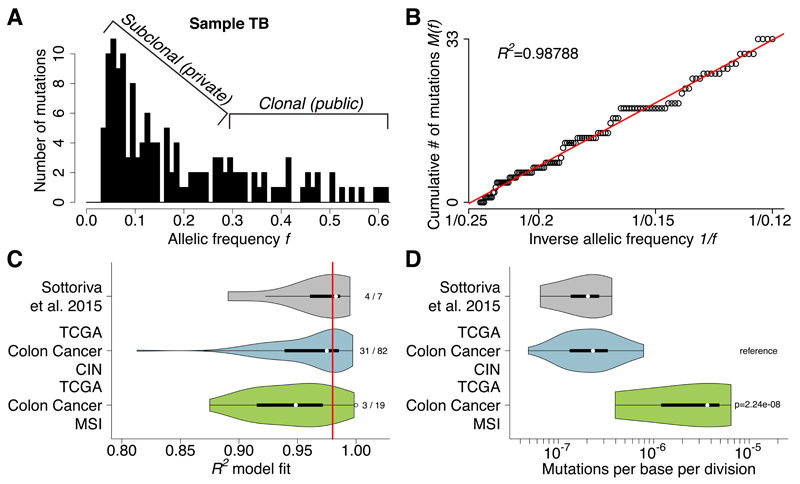
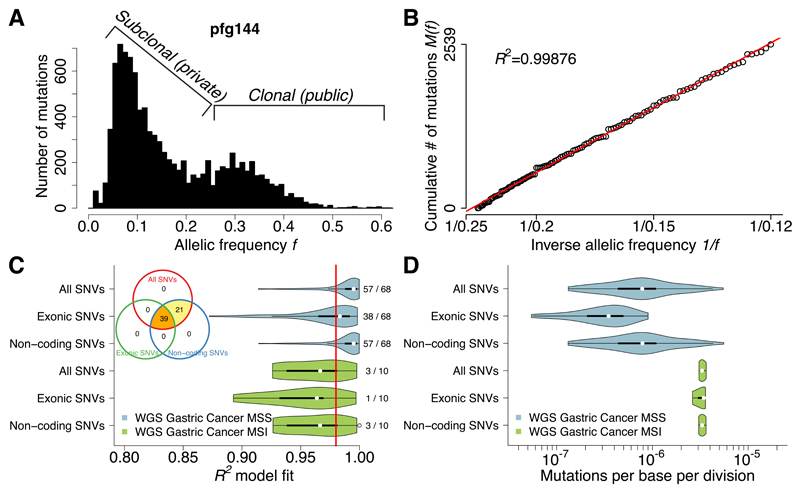
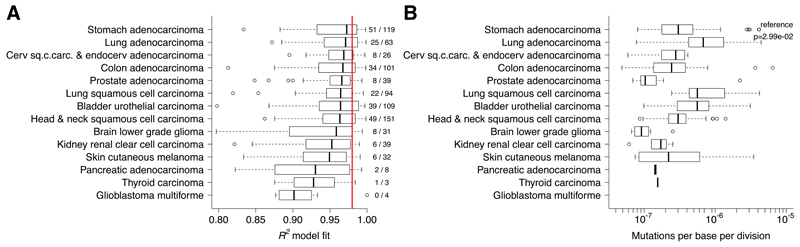
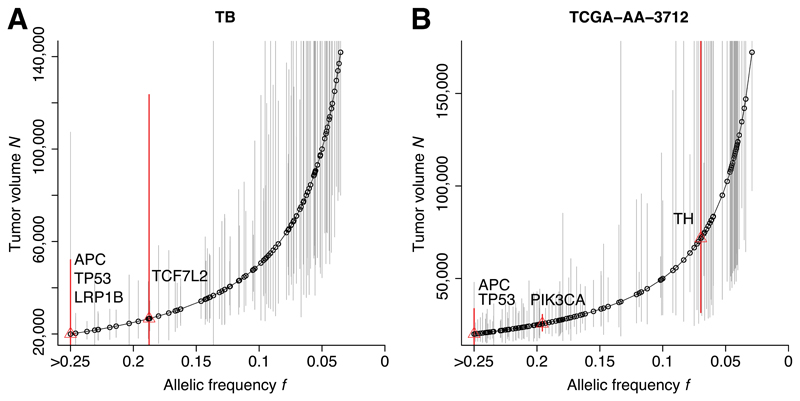
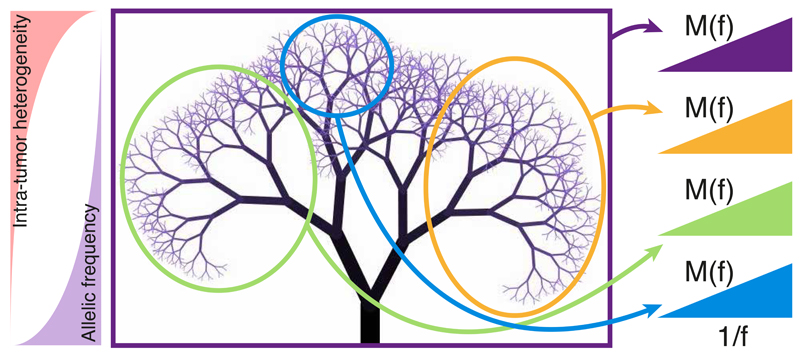
Despite extraordinary efforts to profile cancer genomes, interpreting the vast amount of genomic data in the light of cancer evolution remains challenging. Here we demonstrate that neutral tumor evolution results in a power-law distribution of the mutant allele frequencies reported by next-generation sequencing of tumor bulk samples. We find that the neutral power law fits with high precision 323 of 904 cancers from 14 types and from different cohorts. In malignancies identified as evolving neutrally, all clonal selection seemingly occurred before the onset of cancer growth and not in later-arising subclones, resulting in numerous passenger mutations that are responsible for intratumoral heterogeneity. Reanalyzing cancer sequencing data within the neutral framework allowed the measurement, in each patient, of both the in vivo mutation rate and the order and timing of mutations. This result provides a new way to interpret existing cancer genomic data and to discriminate between functional and non-functional intratumoral heterogeneity.
Figures
Comment in
-
Uncertainties in tumor allele frequencies limit power to infer evolutionary pressures.Nat Genet. 2017 Aug 30;49(9):1288-1289. doi: 10.1038/ng.3876. Nat Genet. 2017. PMID: 28854177 Free PMC article. No abstract available.
-
Revisiting signatures of neutral tumor evolution in the light of complexity of cancer genomic data.Nat Genet. 2018 Dec;50(12):1626-1628. doi: 10.1038/s41588-018-0219-4. Nat Genet. 2018. PMID: 30250123 Free PMC article. No abstract available.
-
Currently available bulk sequencing data do not necessarily support a model of neutral tumor evolution.Nat Genet. 2018 Dec;50(12):1620-1623. doi: 10.1038/s41588-018-0217-6. Nat Genet. 2018. PMID: 30374067 Free PMC article. No abstract available.
-
Neutral tumor evolution?Nat Genet. 2018 Dec;50(12):1630-1633. doi: 10.1038/s41588-018-0258-x. Nat Genet. 2018. PMID: 30374075 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
