

CNS-wide Sexually Dimorphic Induction of the Major Histocompatibility Complex 1 Pathway With Aging
- PMID: 26786204
- PMCID: PMC5155655
- DOI: 10.1093/gerona/glv232
CNS-wide Sexually Dimorphic Induction of the Major Histocompatibility Complex 1 Pathway With Aging
Abstract
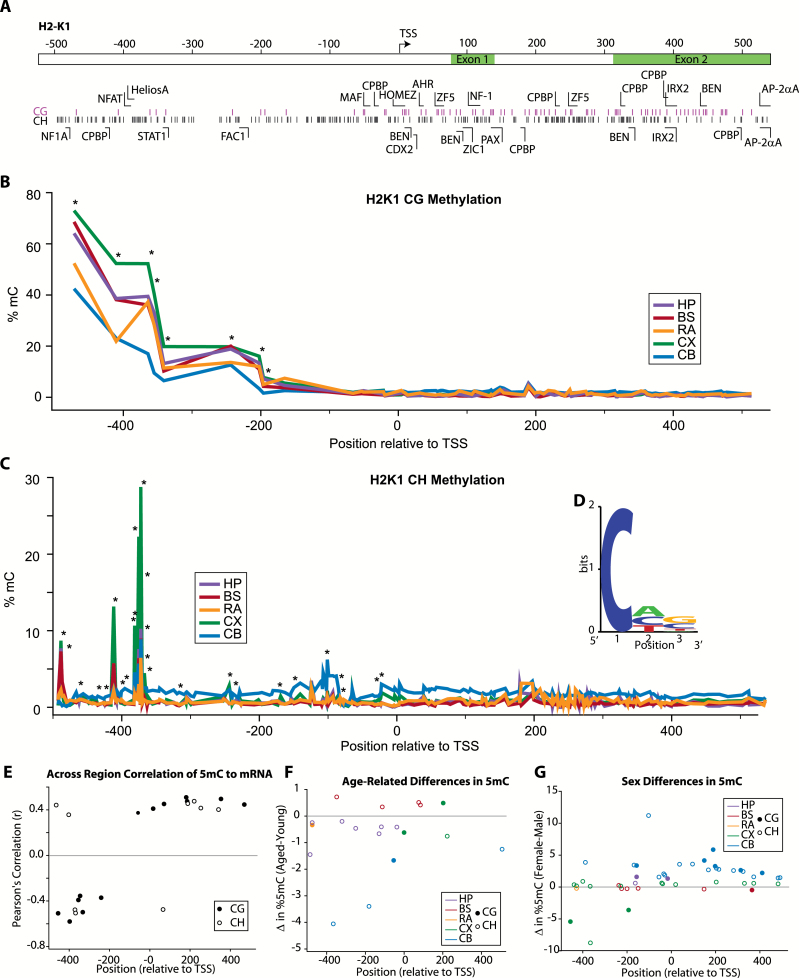
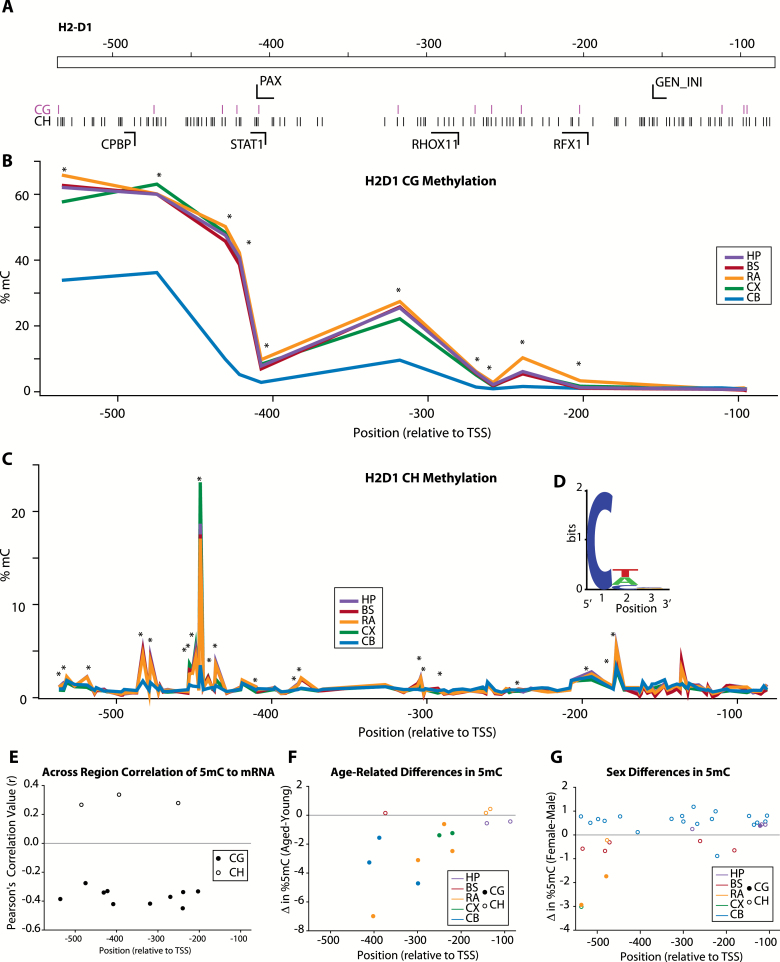
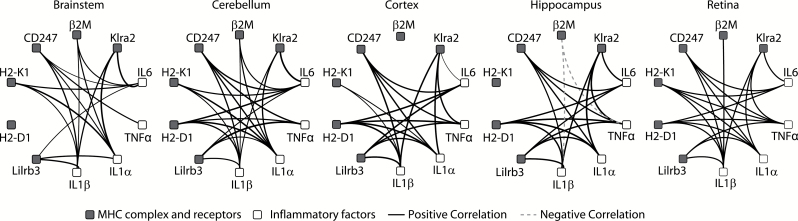
The major histocompatibility complex I (MHCI) pathway, which canonically functions in innate immune viral antigen presentation and detection, is functionally pleiotropic in the central nervous system (CNS). Alternative roles include developmental synapse pruning, regulation of synaptic plasticity, and inhibition of neuronal insulin signaling; all processes altered during brain aging. Upregulation of MHCI components with aging has been reported; however, no systematic examination of MHCI cellular localization, expression, and regulation across CNS regions, life span, and sexes has been reported. In the mouse, MHCI is expressed by neurons and microglia, and MHCI components and receptors (H2-K1, H2-D1, β2M, Lilrb3, Klra2, CD247) display markedly different expression profiles across the hippocampus, cortex, cerebellum, brainstem, and retina. MHCI components, receptors, associated inflammatory transcripts (IL1α, IL1β, IL6, TNFα), and TAP (transporter associated with antigen processing) components are induced with aging and to a greater degree in female than male mice across CNS regions. H2-K1 and H2-D1 expression is associated with differential CG and non-CG promoter methylation across CNS regions, ages, and between sexes, and concomitant increased expression of proinflammatory genes. Meta-analysis of human brain aging data also demonstrates age-related increases in MHCI. Induction of MHCI signaling could contribute to altered synapse regulation and impaired synaptic plasticity with aging.
Keywords: Aging; DNA methylation; Gene expression; MHCI; Sex differences; brain.
© The Author 2016. Published by Oxford University Press on behalf of The Gerontological Society of America. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
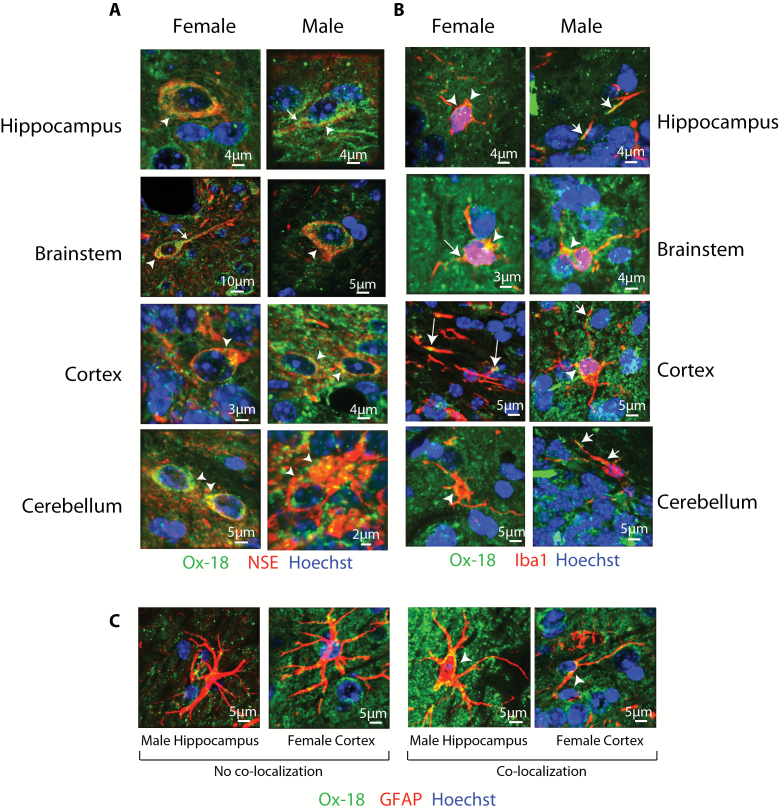
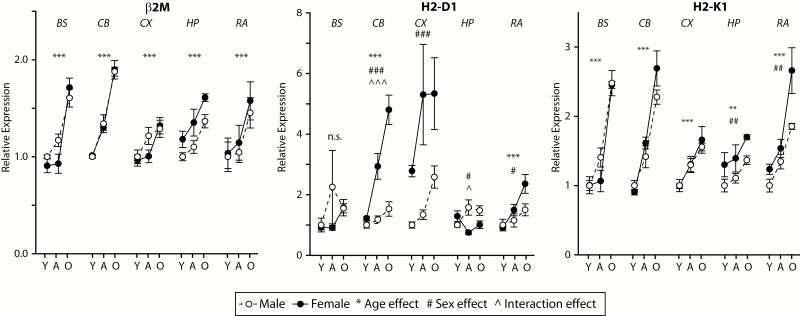
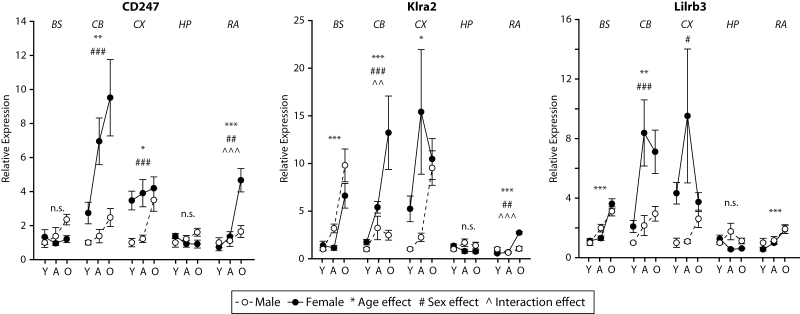
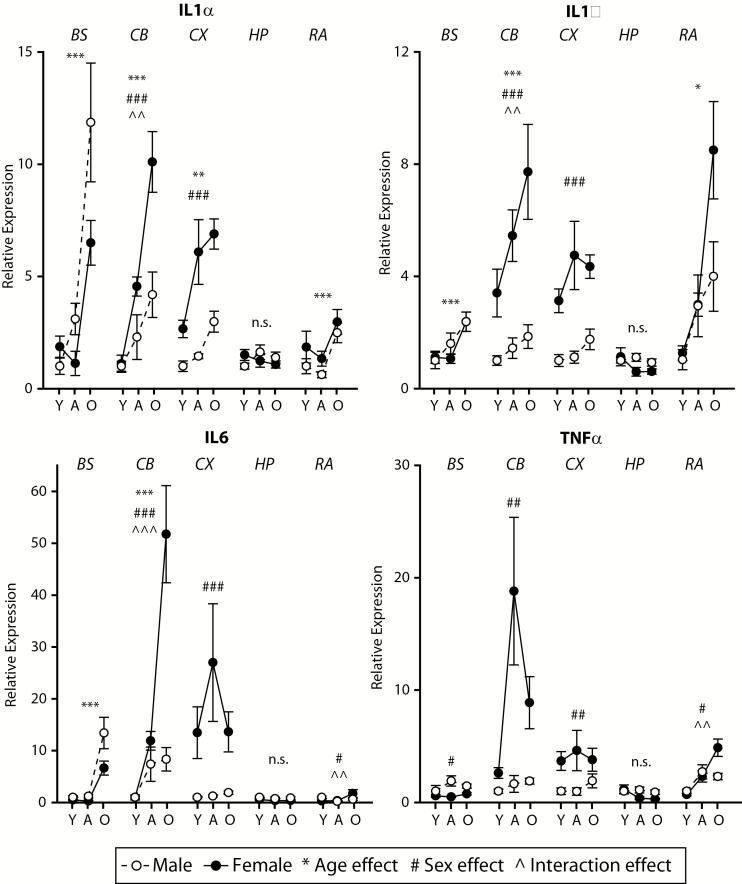
References
-
- Hedden T, Gabrieli JD. Insights into the ageing mind: a view from cognitive neuroscience. Nat Rev Neurosci. 2004;5:87–96. doi:10.1038/nrn1323 - PubMed
-
- Joly E, Mucke L, Oldstone MB. Viral persistence in neurons explained by lack of major histocompatibility class I expression. Science. 1991;253:1283–1285. - PubMed
-
- Neuwelt EA, Clark WK. Unique aspects of central nervous system immunology. Neurosurgery. 1978;3:419–430. - PubMed
-
- Neefjes J, Jongsma ML, Paul P, Bakke O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat Rev Immunol. 2011;11:823–836. doi:10.1038/nri3084 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
