

Pervasive selection for and against antibiotic resistance in inhomogeneous multistress environments
- PMID: 26787239
- PMCID: PMC4735756
- DOI: 10.1038/ncomms10333
Pervasive selection for and against antibiotic resistance in inhomogeneous multistress environments
Abstract
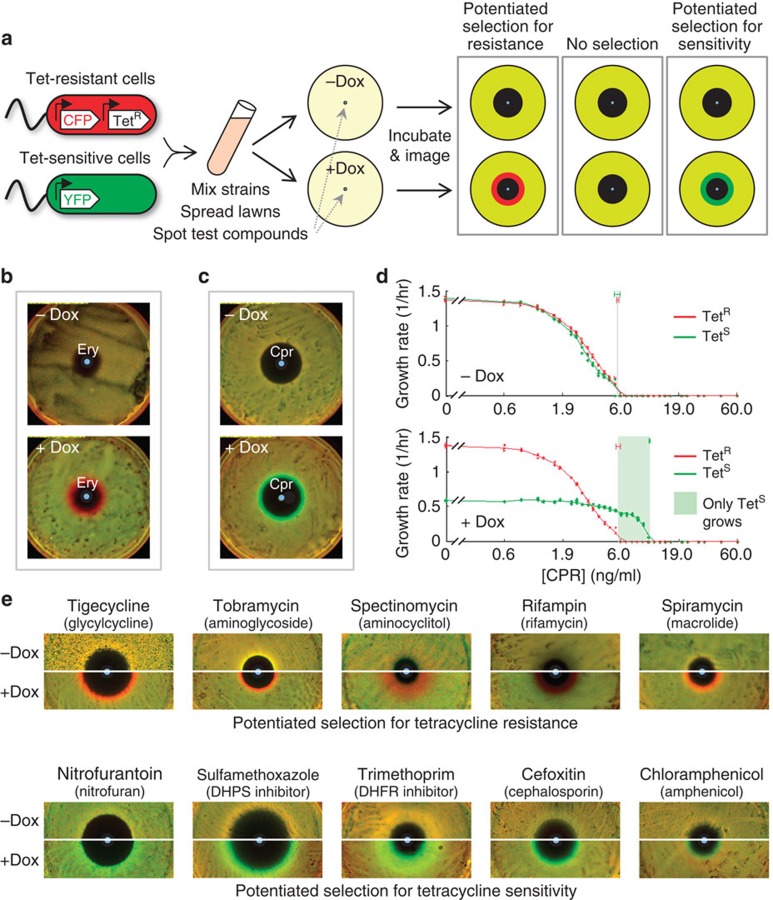
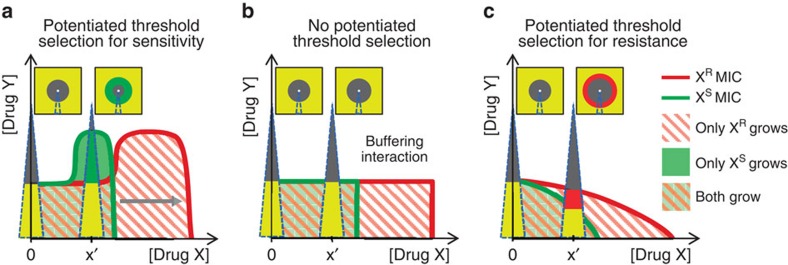
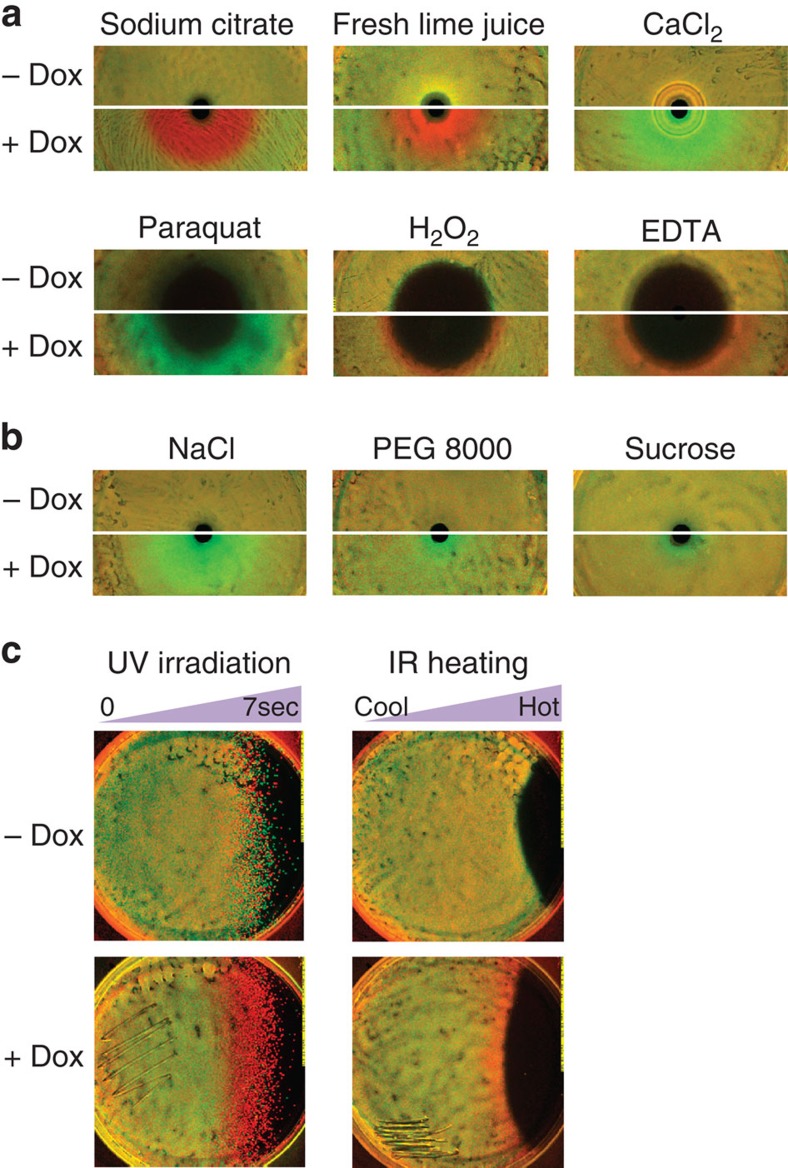
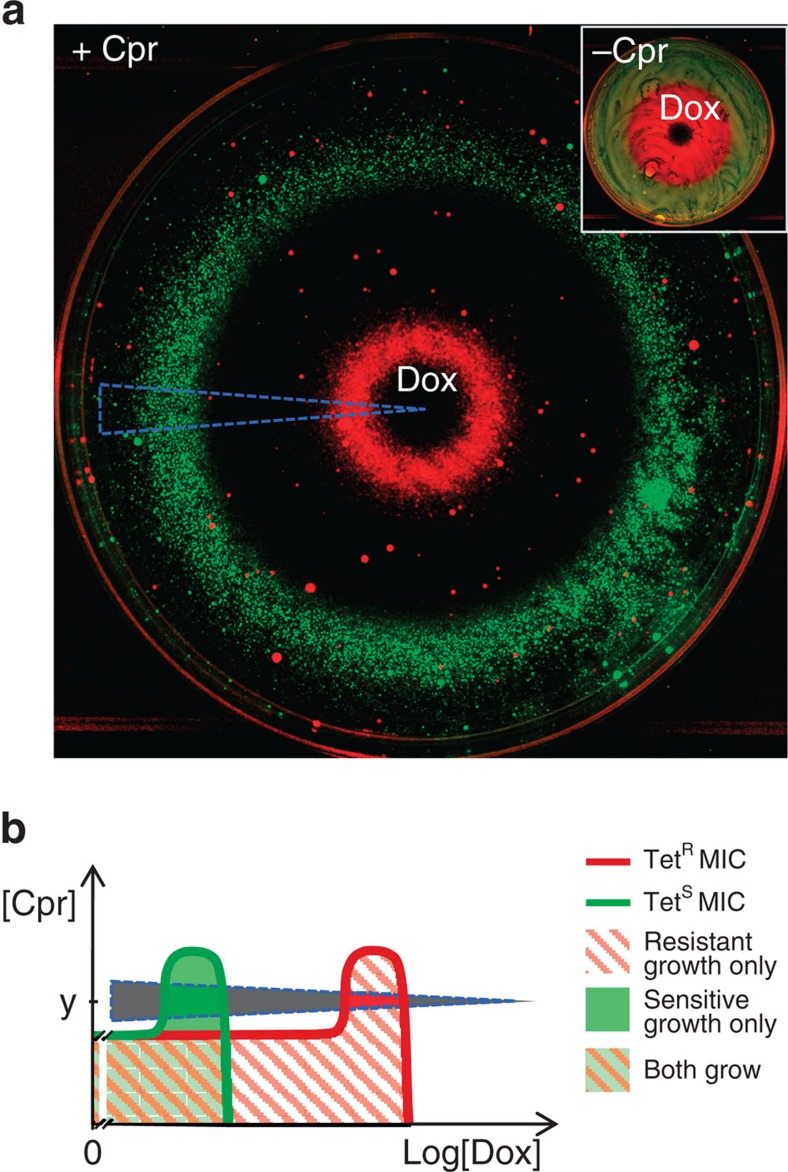
Antibiotic-sensitive and -resistant bacteria coexist in natural environments with low, if detectable, antibiotic concentrations. Except possibly around localized antibiotic sources, where resistance can provide a strong advantage, bacterial fitness is dominated by stresses unaffected by resistance to the antibiotic. How do such mixed and heterogeneous conditions influence the selective advantage or disadvantage of antibiotic resistance? Here we find that sub-inhibitory levels of tetracyclines potentiate selection for or against tetracycline resistance around localized sources of almost any toxin or stress. Furthermore, certain stresses generate alternating rings of selection for and against resistance around a localized source of the antibiotic. In these conditions, localized antibiotic sources, even at high strengths, can actually produce a net selection against resistance to the antibiotic. Our results show that interactions between the effects of an antibiotic and other stresses in inhomogeneous environments can generate pervasive, complex patterns of selection both for and against antibiotic resistance.
Figures
Similar articles
-
Antibiotic interactions that select against resistance.Nature. 2007 Apr 5;446(7136):668-71. doi: 10.1038/nature05685. Nature. 2007. PMID: 17410176
-
Organic acids enhance bioavailability of tetracycline in water to Escherichia coli for uptake and expression of antibiotic resistance.Water Res. 2014 Nov 15;65:98-106. doi: 10.1016/j.watres.2014.07.021. Epub 2014 Jul 22. Water Res. 2014. PMID: 25100186
-
Impact of Cross-Contamination Concentrations of Doxycycline Hyclate on the Microbial Ecosystem in an Ex Vivo Model of the Pig's Cecum.Microb Drug Resist. 2019 Mar;25(2):304-315. doi: 10.1089/mdr.2018.0034. Epub 2018 Sep 19. Microb Drug Resist. 2019. PMID: 30234420
-
Antibiotic-selective environments.Clin Infect Dis. 1998 Aug;27 Suppl 1:S5-11. doi: 10.1086/514916. Clin Infect Dis. 1998. PMID: 9710666 Review.
-
[Genetic and biochemical basis of microbial resistance to antibiotics].Lijec Vjesn. 1990 Jul-Aug;112(7-8):250-7. Lijec Vjesn. 1990. PMID: 2292900 Review. Croatian.
Cited by
-
Highly Contingent Phenotypes of Lon Protease Deficiency in Escherichia coli upon Antibiotic Challenge.J Bacteriol. 2020 Jan 15;202(3):e00561-19. doi: 10.1128/JB.00561-19. Print 2020 Jan 15. J Bacteriol. 2020. PMID: 31740490 Free PMC article.
-
Efflux pump-mediated resistance to antifungal compounds can be prevented by conjugation with triphenylphosphonium cation.Nat Commun. 2018 Nov 30;9(1):5102. doi: 10.1038/s41467-018-07633-9. Nat Commun. 2018. PMID: 30504815 Free PMC article.
-
Engineered bacteria for near-infrared light-inducible expression of cancer therapeutics.Nat Cancer. 2025 Apr;6(4):612-628. doi: 10.1038/s43018-025-00932-3. Epub 2025 Mar 17. Nat Cancer. 2025. PMID: 40097656
-
The Persistence of Antibiotic Resistance in Observational Studies: Is It Really Due to Differences in Sub-Populations Rather than Antibiotic Use?Antibiotics (Basel). 2025 Jan 6;14(1):39. doi: 10.3390/antibiotics14010039. Antibiotics (Basel). 2025. PMID: 39858325 Free PMC article.
-
Prevalence of antibiotic resistance genes its association with microbiota in raw milk of northwest Xinjiang.Front Microbiol. 2025 Jul 11;16:1595051. doi: 10.3389/fmicb.2025.1595051. eCollection 2025. Front Microbiol. 2025. PMID: 40718809 Free PMC article.
References
-
- Levy S. B. & Marshall B. Antibacterial resistance worldwide: causes, challenges and responses. Nat. Med. 10, S122–S129 (2004) . - PubMed
-
- D'Costa V. M., McGrann K. M., Hughes D. W. & Wright G. D. Sampling the antibiotic resistome. Science 311, 374–377 (2006) . - PubMed
-
- Waksman S. A. The role of antibiotics in nature. Perspect. Biol. Med. 4, 271–287 (1961) .
-
- Gottlieb D. The production and role of antibiotics in soil. J. Antibiot. (Tokyo) 29, 987–1000 (1976) . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
