

Nuclear hormone receptor DHR96 mediates the resistance to xenobiotics but not the increased lifespan of insulin-mutant Drosophila
- PMID: 26787908
- PMCID: PMC4747718
- DOI: 10.1073/pnas.1515137113
Nuclear hormone receptor DHR96 mediates the resistance to xenobiotics but not the increased lifespan of insulin-mutant Drosophila
Abstract
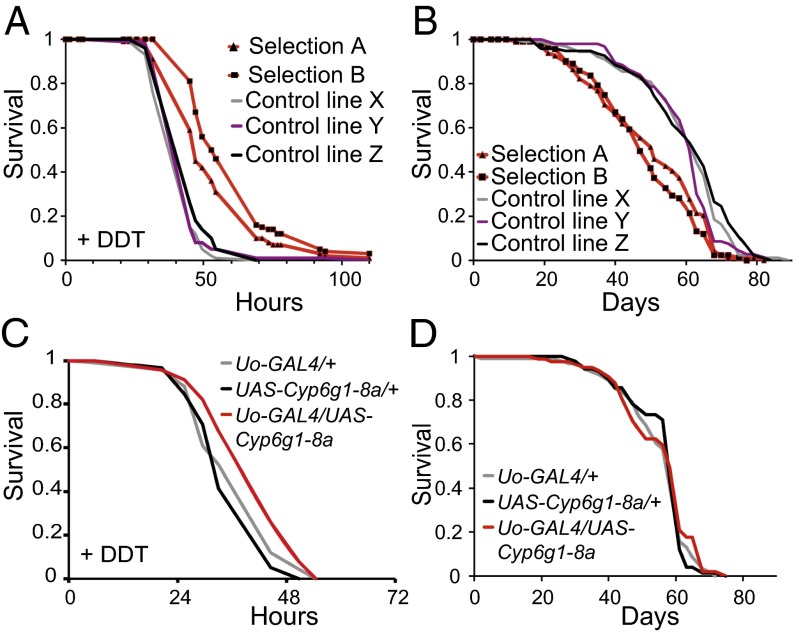
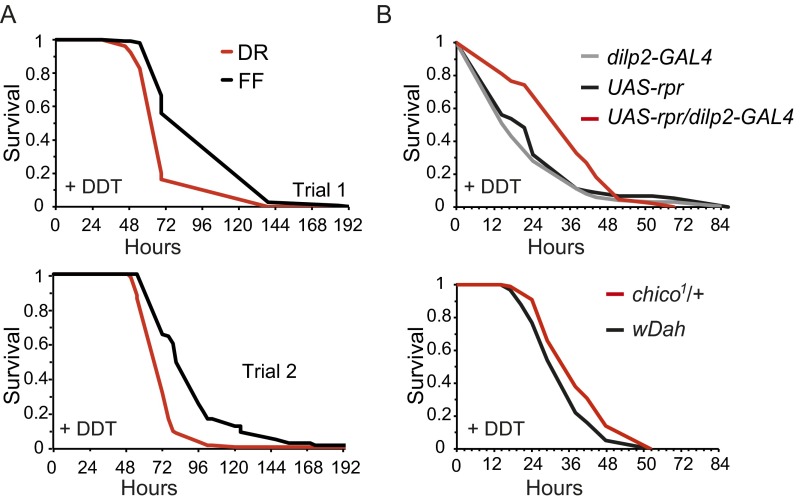
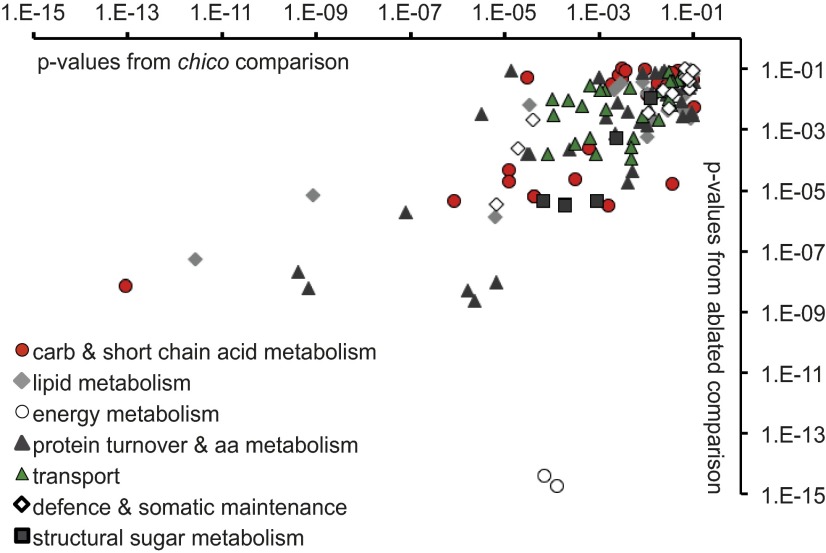
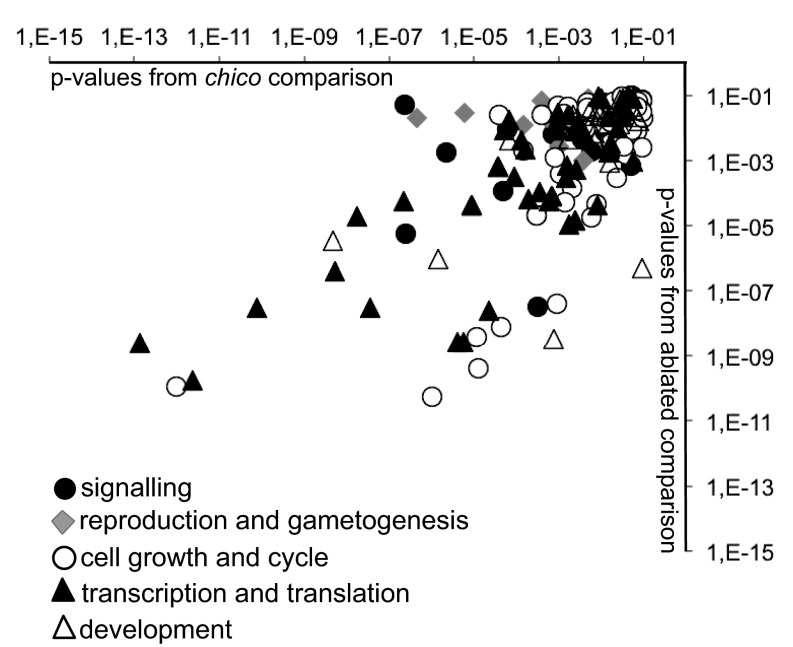
Lifespan of laboratory animals can be increased by genetic, pharmacological, and dietary interventions. Increased expression of genes involved in xenobiotic metabolism, together with resistance to xenobiotics, are frequent correlates of lifespan extension in the nematode worm Caenorhabditis elegans, the fruit fly Drosophila, and mice. The Green Theory of Aging suggests that this association is causal, with the ability of cells to rid themselves of lipophilic toxins limiting normal lifespan. To test this idea, we experimentally increased resistance of Drosophila to the xenobiotic dichlordiphenyltrichlorethan (DDT), by artificial selection or by transgenic expression of a gene encoding a cytochrome P450. Although both interventions increased DDT resistance, neither increased lifespan. Furthermore, dietary restriction increased lifespan without increasing xenobiotic resistance, confirming that the two traits can be uncoupled. Reduced activity of the insulin/Igf signaling (IIS) pathway increases resistance to xenobiotics and extends lifespan in Drosophila, and can also increase longevity in C. elegans, mice, and possibly humans. We identified a nuclear hormone receptor, DHR96, as an essential mediator of the increased xenobiotic resistance of IIS mutant flies. However, the IIS mutants remained long-lived in the absence of DHR96 and the xenobiotic resistance that it conferred. Thus, in Drosophila IIS mutants, increased xenobiotic resistance and enhanced longevity are not causally connected. The frequent co-occurrence of the two traits may instead have evolved because, in nature, lowered IIS can signal the presence of pathogens. It will be important to determine whether enhanced xenobiotic metabolism is also a correlated, rather than a causal, trait in long-lived mice.
Keywords: DHR96; IIS; lifespan; nuclear hormone receptor; xenobiotic resistance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
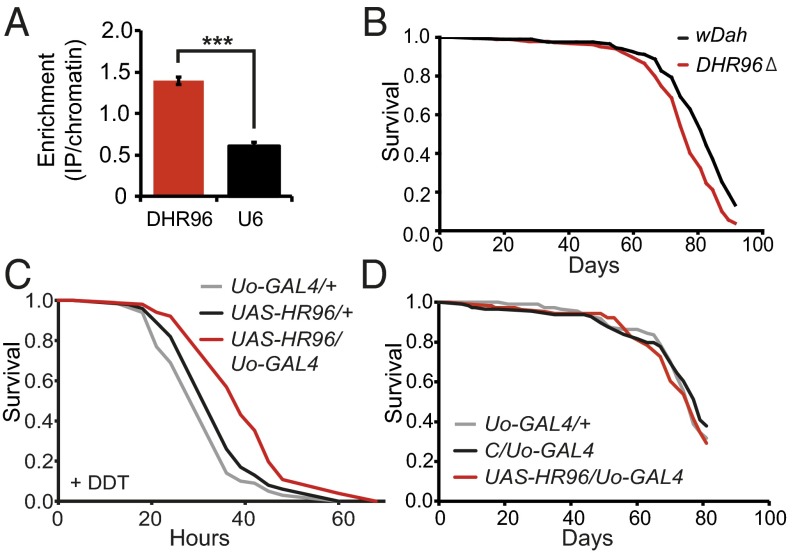
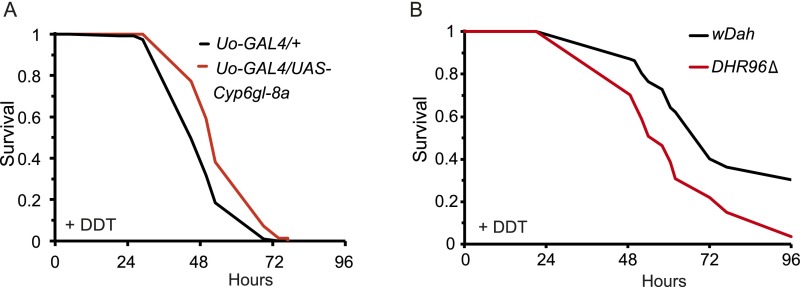

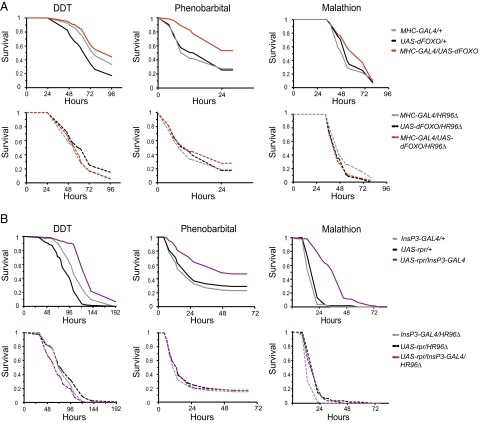
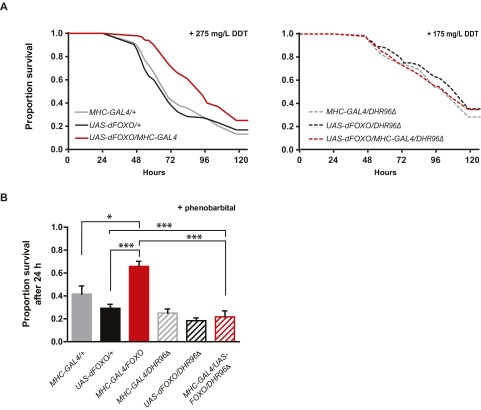
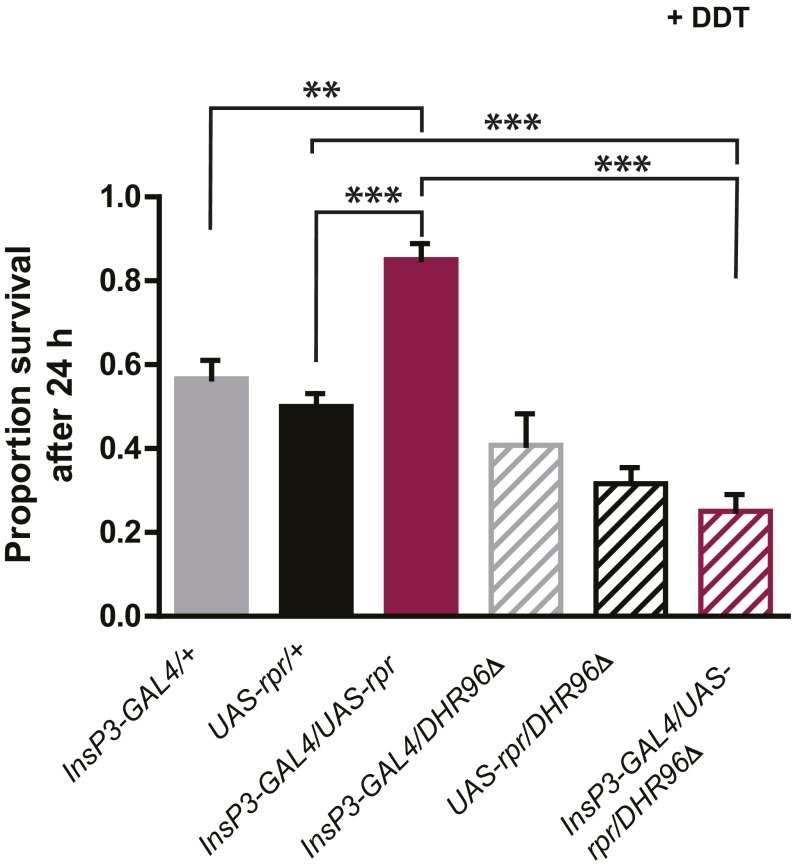
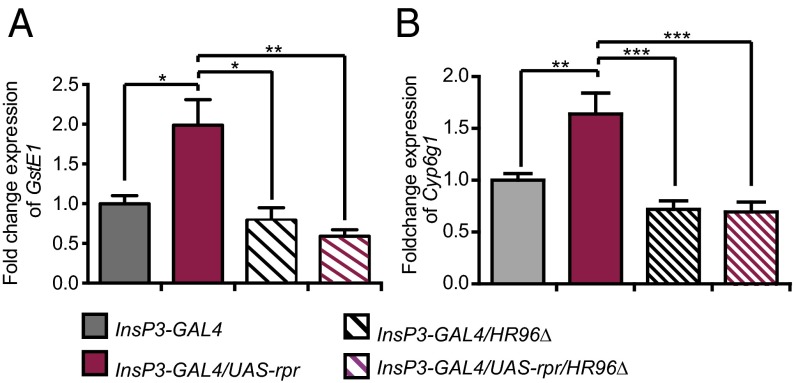
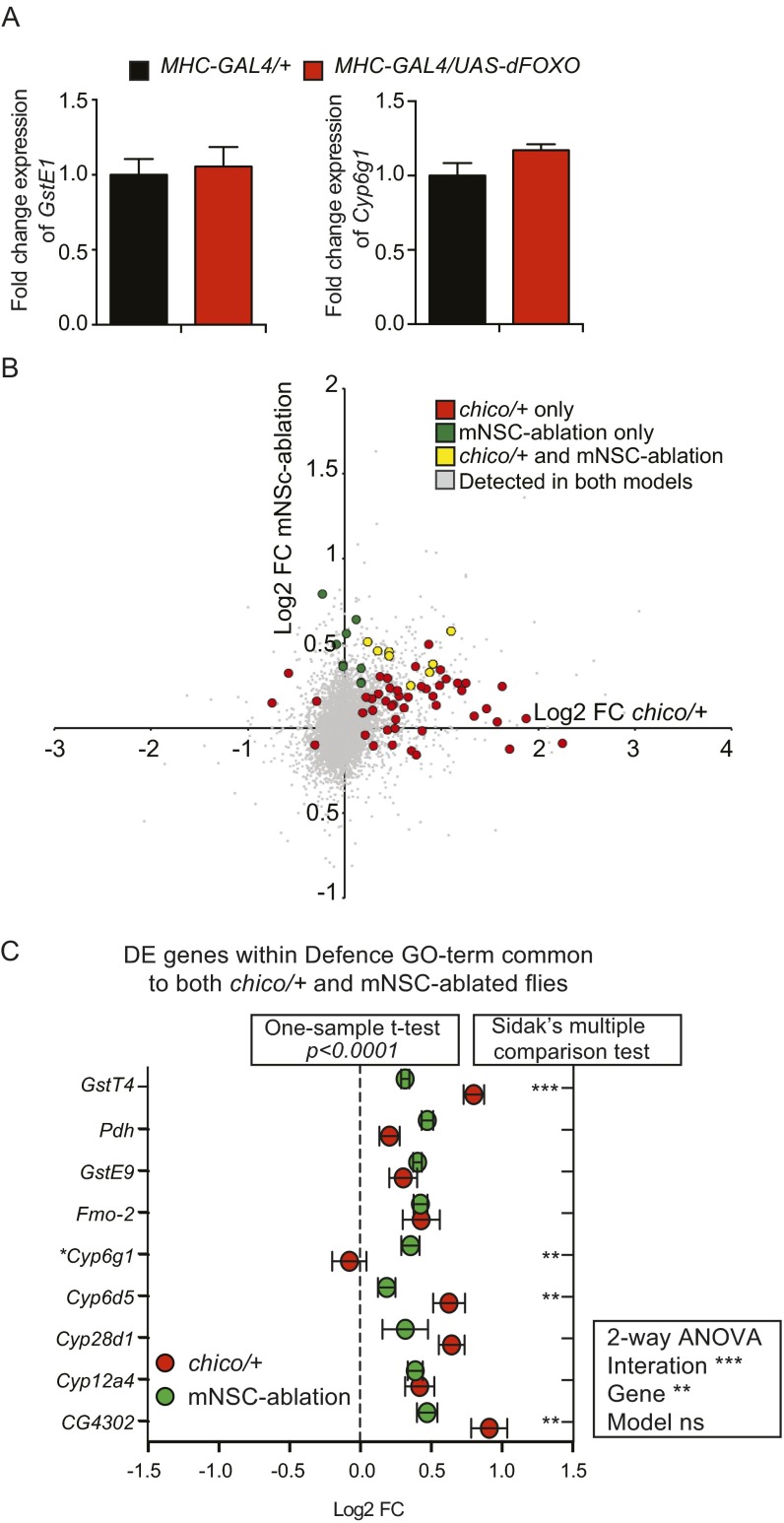
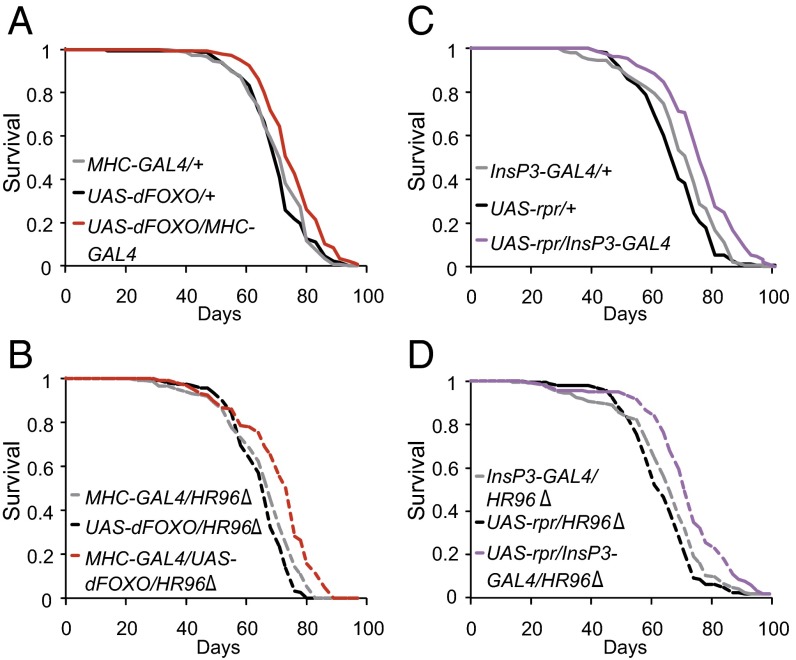
References
-
- Kenyon CJ. The genetics of ageing. Nature. 2010;464(7288):504–512. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
