

The Behavioral Relevance of Cortical Neural Ensemble Responses Emerges Suddenly
- PMID: 26791199
- PMCID: PMC4719008
- DOI: 10.1523/JNEUROSCI.2265-15.2016
The Behavioral Relevance of Cortical Neural Ensemble Responses Emerges Suddenly
Abstract
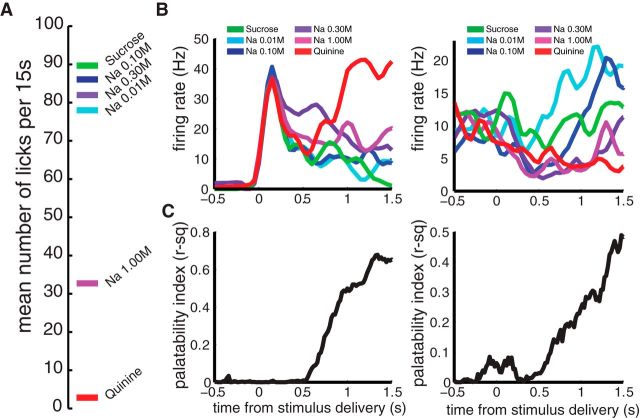
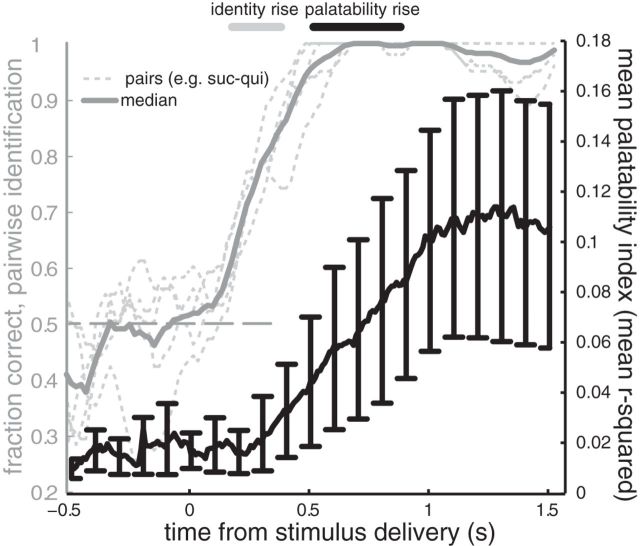
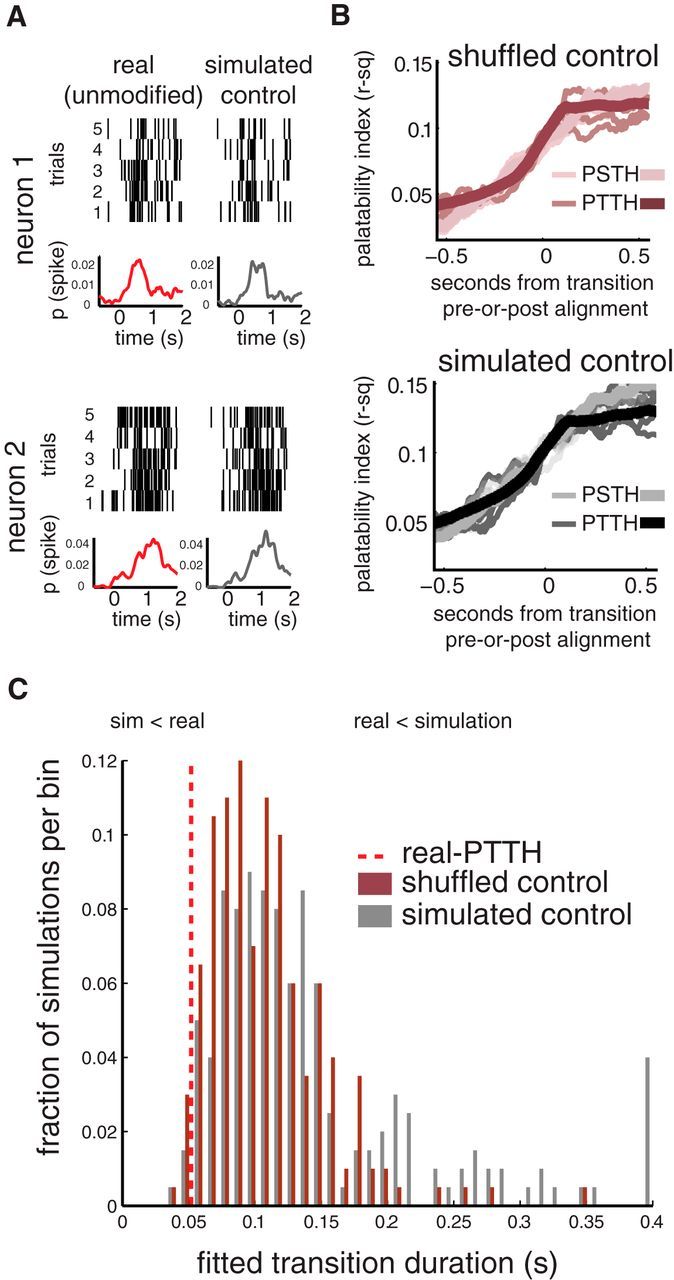
Whereas many laboratory-studied decisions involve a highly trained animal identifying an ambiguous stimulus, many naturalistic decisions do not. Consumption decisions, for instance, involve determining whether to eject or consume an already identified stimulus in the mouth and are decisions that can be made without training. By standard analyses, rodent cortical single-neuron taste responses come to predict such consumption decisions across the 500 ms preceding the consumption or rejection itself; decision-related firing emerges well after stimulus identification. Analyzing single-trial ensemble activity using hidden Markov models, we show these decision-related cortical responses to be part of a reliable sequence of states (each defined by the firing rates within the ensemble) separated by brief state-to-state transitions, the latencies of which vary widely between trials. When we aligned data to the onset of the (late-appearing) state that dominates during the time period in which single-neuron firing is correlated to taste palatability, the apparent ramp in stimulus-aligned choice-related firing was shown to be a much more precipitous coherent jump. This jump in choice-related firing resembled a step function more than it did the output of a standard (ramping) decision-making model, and provided a robust prediction of decision latency in single trials. Together, these results demonstrate that activity related to naturalistic consumption decisions emerges nearly instantaneously in cortical ensembles. Significance statement: This paper provides a description of how the brain makes evaluative decisions. The majority of work on the neurobiology of decision making deals with "what is it?" decisions; out of this work has emerged a model whereby neurons accumulate information about the stimulus in the form of slowly increasing firing rates and reach a decision when those firing rates reach a threshold. Here, we study a different kind of more naturalistic decision--a decision to evaluate "what shall I do with it?" after the identity of a taste in the mouth has been identified--and show that this decision is not made through the gradual increasing of stimulus-related firing, but rather that this decision appears to be made in a sudden moment of "insight."
Keywords: attractor; cortex; decision; gustatory; hidden Markov models; taste.
Copyright © 2016 the authors 0270-6474/16/360655-15$15.00/0.
Figures




References
-
- Baum LE, Petrie T, Soules G, Weiss N. A maximization technique occurring in the statistical analysis of probabilistic functions of Markov chains. Ann Math Stat. 1970;41:164–171. doi: 10.1214/aoms/1177697196. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical