

Short Fibrils Constitute the Major Species of Seed-Competent Tau in the Brains of Mice Transgenic for Human P301S Tau
- PMID: 26791207
- PMCID: PMC4719013
- DOI: 10.1523/JNEUROSCI.3542-15.2016
Short Fibrils Constitute the Major Species of Seed-Competent Tau in the Brains of Mice Transgenic for Human P301S Tau
Abstract
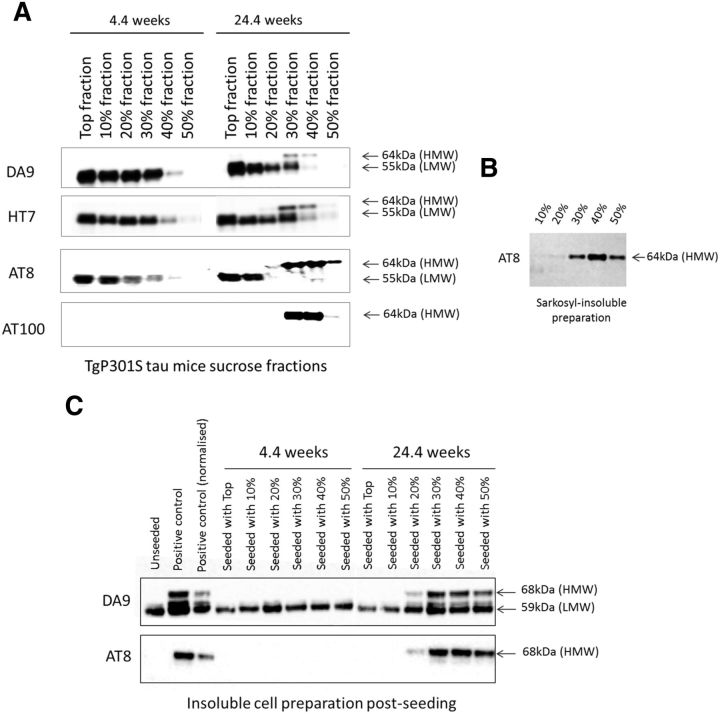
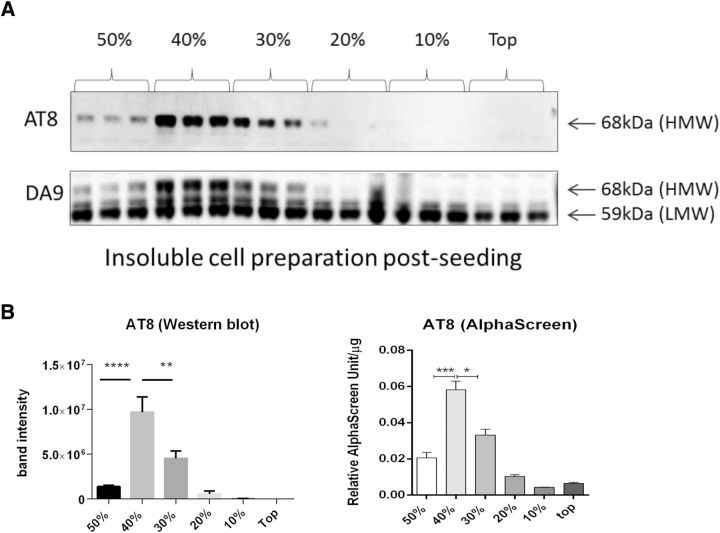
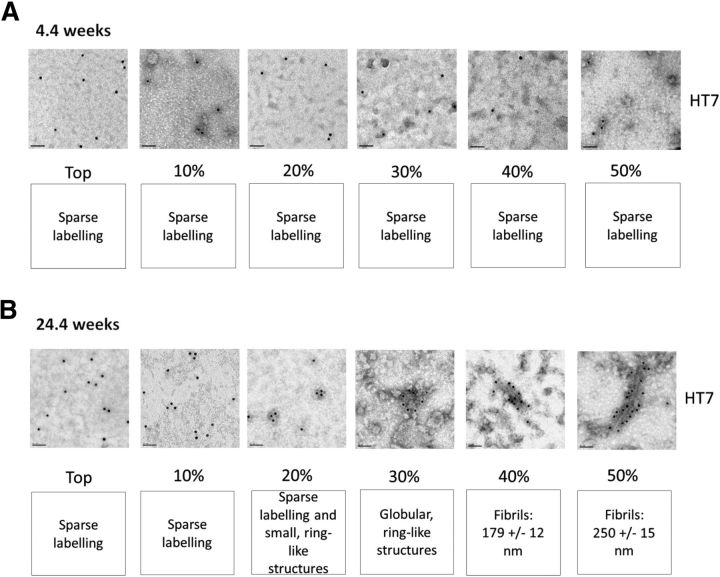
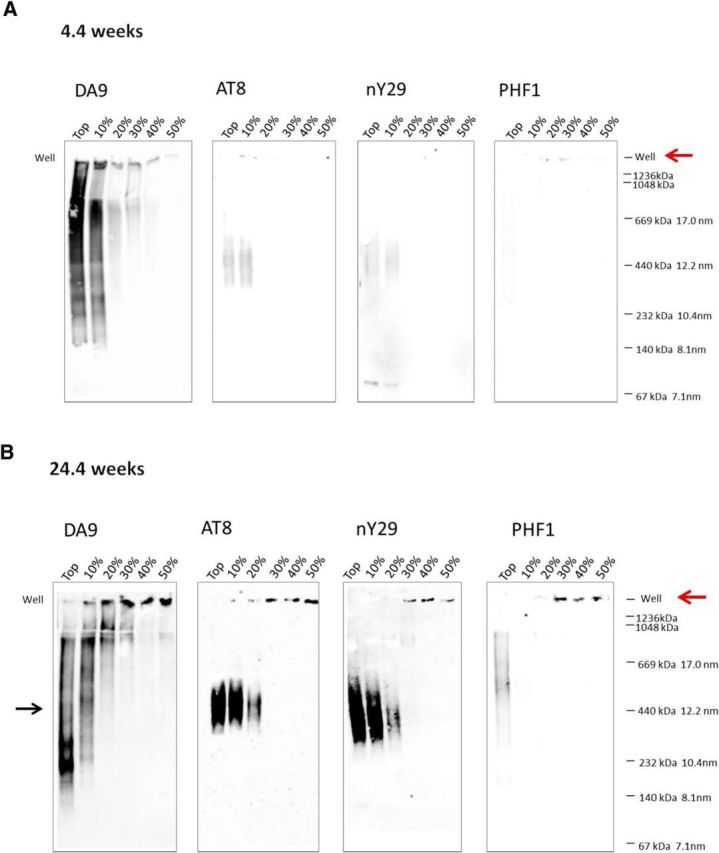
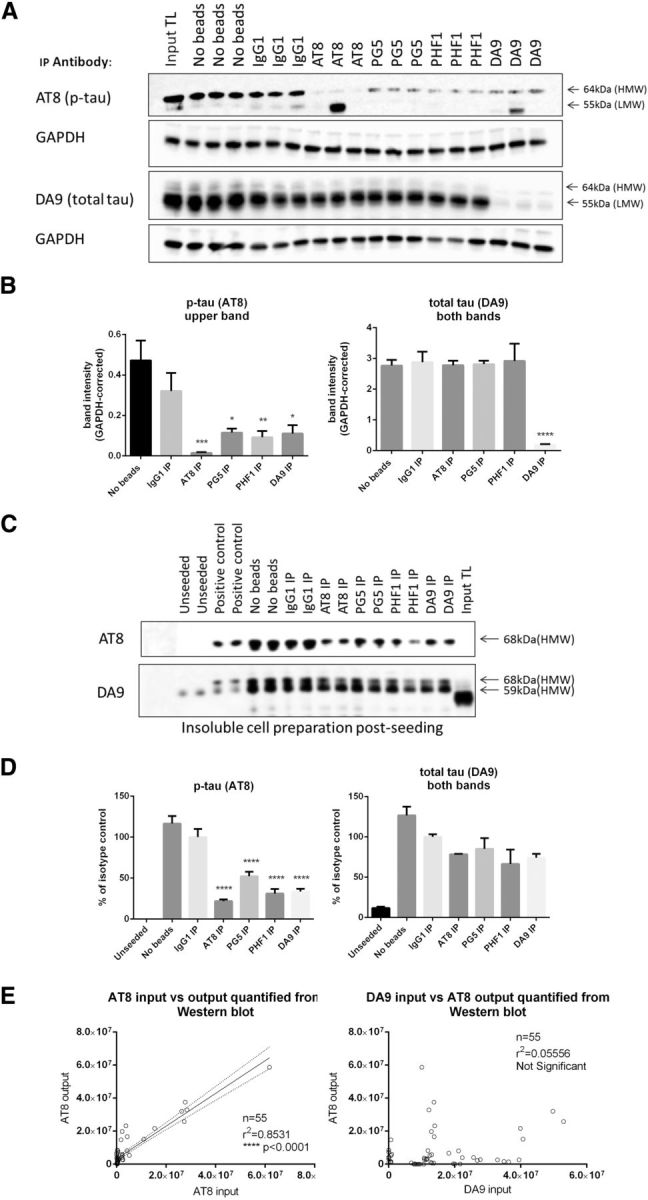
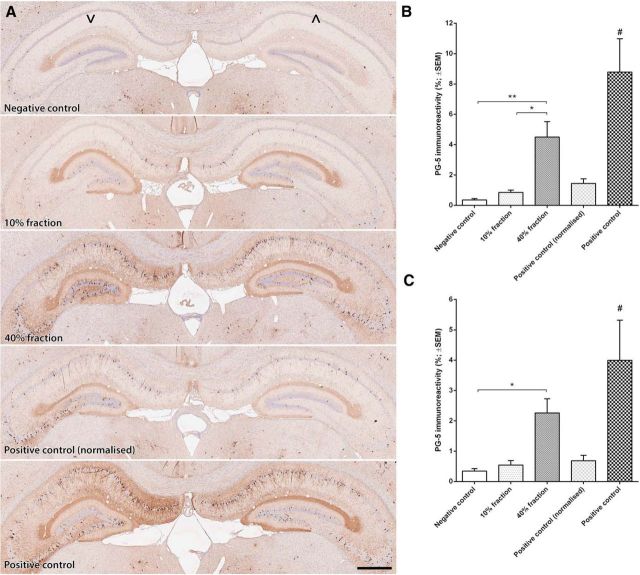
The interneuronal propagation of aggregated tau is believed to play an important role in the pathogenesis of human tauopathies. It requires the uptake of seed-competent tau into cells, seeding of soluble tau in recipient neurons and release of seeded tau into the extracellular space to complete the cycle. At present, it is not known which tau species are seed-competent. Here, we have dissected the molecular characteristics of seed-competent tau species from the TgP301S tau mouse model using various biochemical techniques and assessed their seeding ability in cell and animal models. We found that sucrose gradient fractions from brain lysates seeded cellular tau aggregation only when large (>10 mer) aggregated, hyperphosphorylated (AT8- and AT100-positive) and nitrated tau was present. In contrast, there was no detectable seeding by fractions containing small, oligomeric (<6 mer) tau. Immunodepletion of the large aggregated AT8-positive tau strongly reduced seeding; moreover, fractions containing these species initiated the formation and spreading of filamentous tau pathology in vivo, whereas fractions containing tau monomers and small oligomeric assemblies did not. By electron microscopy, seed-competent sucrose gradient fractions contained aggregated tau species ranging from ring-like structures to small filaments. Together, these findings indicate that a range of filamentous tau aggregates are the major species that underlie the spreading of tau pathology in the P301S transgenic model. Significance statement: The spread of tau pathology from neuron to neuron is postulated to account for, or at least to contribute to, the overall propagation of tau pathology during the development of human tauopathies including Alzheimer's disease. It is therefore important to characterize the native tau species responsible for this process of seeding and pathology spreading. Here, we use several biochemical techniques to dissect the molecular characteristics of native tau protein conformers from TgP301S tau mice and show that seed-competent tau species comprise small fibrils capable of seeding tau pathology in cell and animal models. Characterization of seed-competent tau gives insight into disease mechanisms and therapeutic interventions.
Keywords: aggregation; propagation; tau; tauopathy.
Copyright © 2016 Jackson, Kerridge et al.
Figures
References
-
- Ahmed Z, Cooper J, Murray TK, Garn K, McNaughton E, Clarke H, Parhizkar S, Ward MA, Cavallini A, Jackson S, Bose S, Clavaguera F, Tolnay M, Lavenir I, Goedert M, Hutton ML, O'Neill MJ. A novel in vivo model of tau propagation with rapid and progressive neurofibrillary tangle pathology: the pattern of spread is determined by connectivity, not proximity. Acta Neuropathol. 2014;127:667–683. doi: 10.1007/s00401-014-1254-6. - DOI - PMC - PubMed
-
- Allen B, Ingram E, Takao M, Smith MJ, Jakes R, Virdee K, Yoshida H, Holzer M, Craxton M, Emson PC, Atzori C, Migheli A, Crowther RA, Ghetti B, Spillantini MG, Goedert M. Abundant tau filaments and nonapoptotic neurodegeneration in transgenic mice expressing human P301S tau protein. J Neurosci. 2002;22:9340–9351. - PMC - PubMed
-
- Berger Z, Roder H, Hanna A, Carlson A, Rangachari V, Yue M, Wszolek Z, Ashe K, Knight J, Dickson D, Andorfer C, Rosenberry TL, Lewis J, Hutton M, Janus C. Accumulation of pathological tau species and memory loss in a conditional model of tauopathy. J Neurosci. 2007;27:3650–3662. doi: 10.1523/JNEUROSCI.0587-07.2007. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous