

Meiotic recombination counteracts male-biased mutation (male-driven evolution)
- PMID: 26791621
- PMCID: PMC4795034
- DOI: 10.1098/rspb.2015.2691
Meiotic recombination counteracts male-biased mutation (male-driven evolution)
Abstract
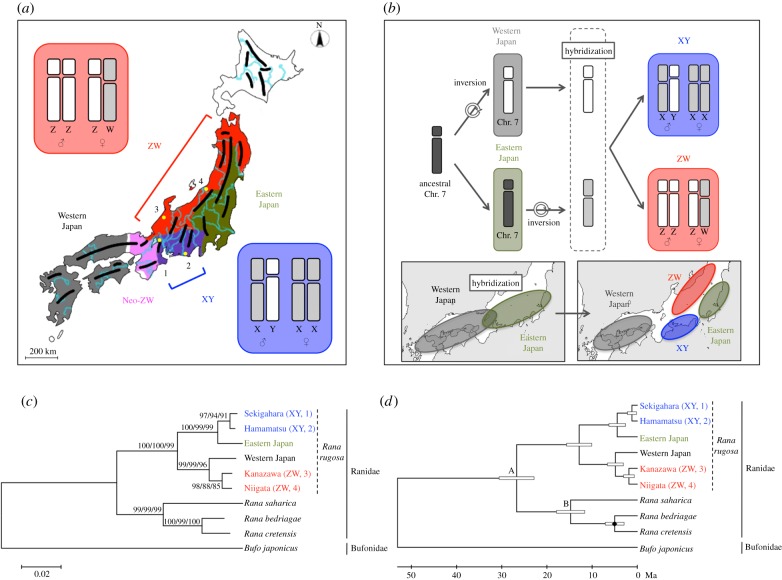
Meiotic recombination is believed to produce greater genetic variation despite the fact that deoxyribonucleic acid (DNA)-replication errors are a major source of mutations. In some vertebrates, mutation rates are higher in males than in females, which developed the theory of male-driven evolution (male-biased mutation). However, there is little molecular evidence regarding the relationships between meiotic recombination and male-biased mutation. Here we tested the theory using the frog Rana rugosa, which has both XX/XY- and ZZ/ZW-type sex-determining systems within the species. The male-to-female mutation-rate ratio (α) was calculated from homologous sequences on the X/Y or Z/W sex chromosomes, which supported male-driven evolution. Surprisingly, each α value was notably higher in the XX/XY-type group than in the ZZ/ZW-type group, although α should have similar values within a species. Interestingly, meiotic recombination between homologous chromosomes did not occur except at terminal regions in males of this species. Then, by subdividing α into two new factors, a replication-based male-to-female mutation-rate ratio (β) and a meiotic recombination-based XX-to-XY/ZZ-to-ZW mutation-rate ratio (γ), we constructed a formula describing the relationship among a nucleotide-substitution rate and the two factors, β and γ. Intriguingly, the β- and γ-values were larger and smaller than 1, respectively, indicating that meiotic recombination might reduce male-biased mutations.
Keywords: germ cell; male-biased mutation; male-driven evolution; meiotic recombination; mutation rate; sex chromosome.
© 2016 The Author(s).
Figures


References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
