

Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures
- PMID: 26792235
- PMCID: PMC4726131
- DOI: 10.1038/srep19715
Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures
Erratum in
-
Corrigendum: Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures.Sci Rep. 2016 May 11;6:25284. doi: 10.1038/srep25284. Sci Rep. 2016. PMID: 27166264 Free PMC article. No abstract available.
Abstract
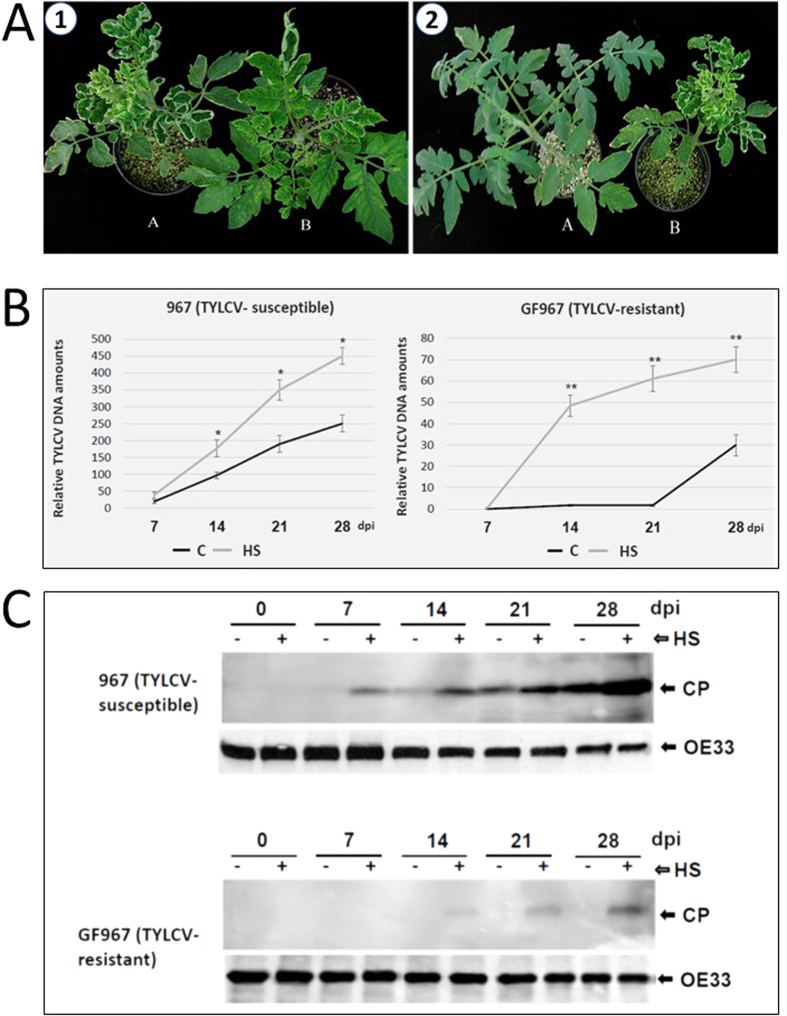
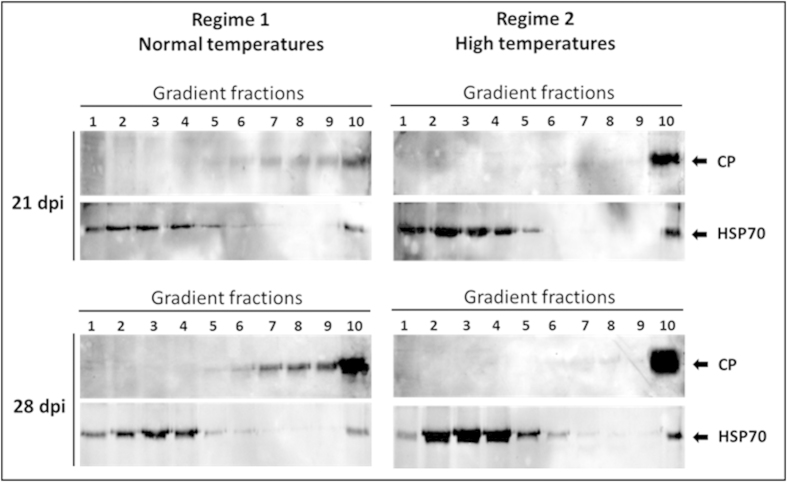
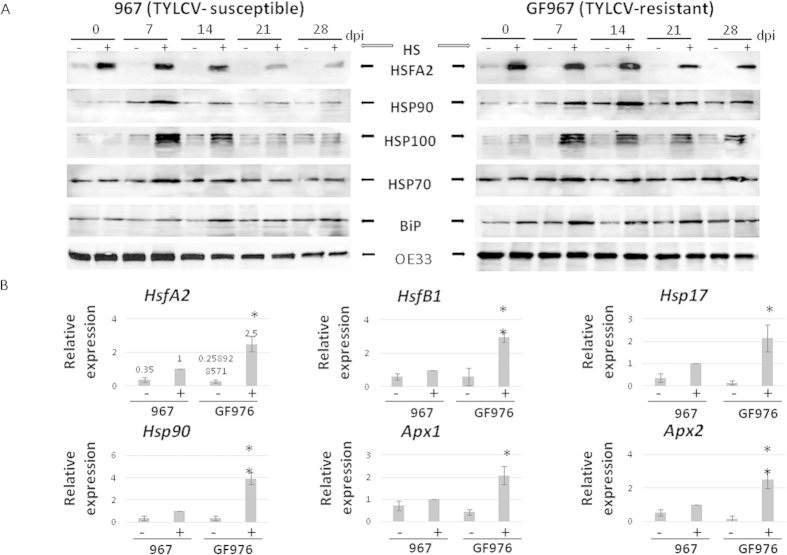
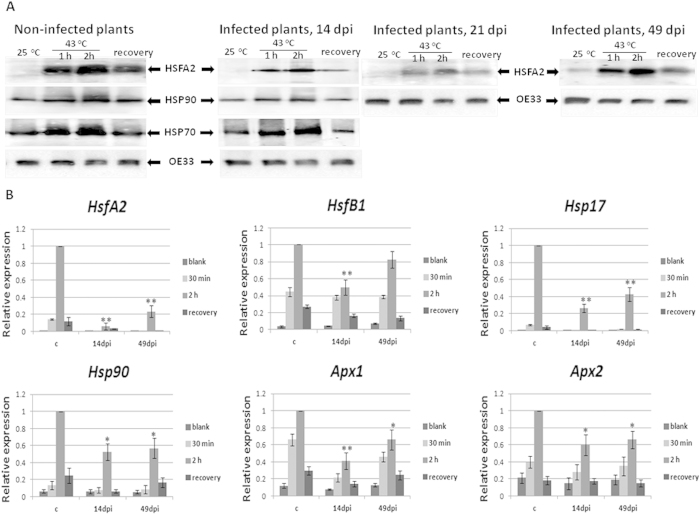
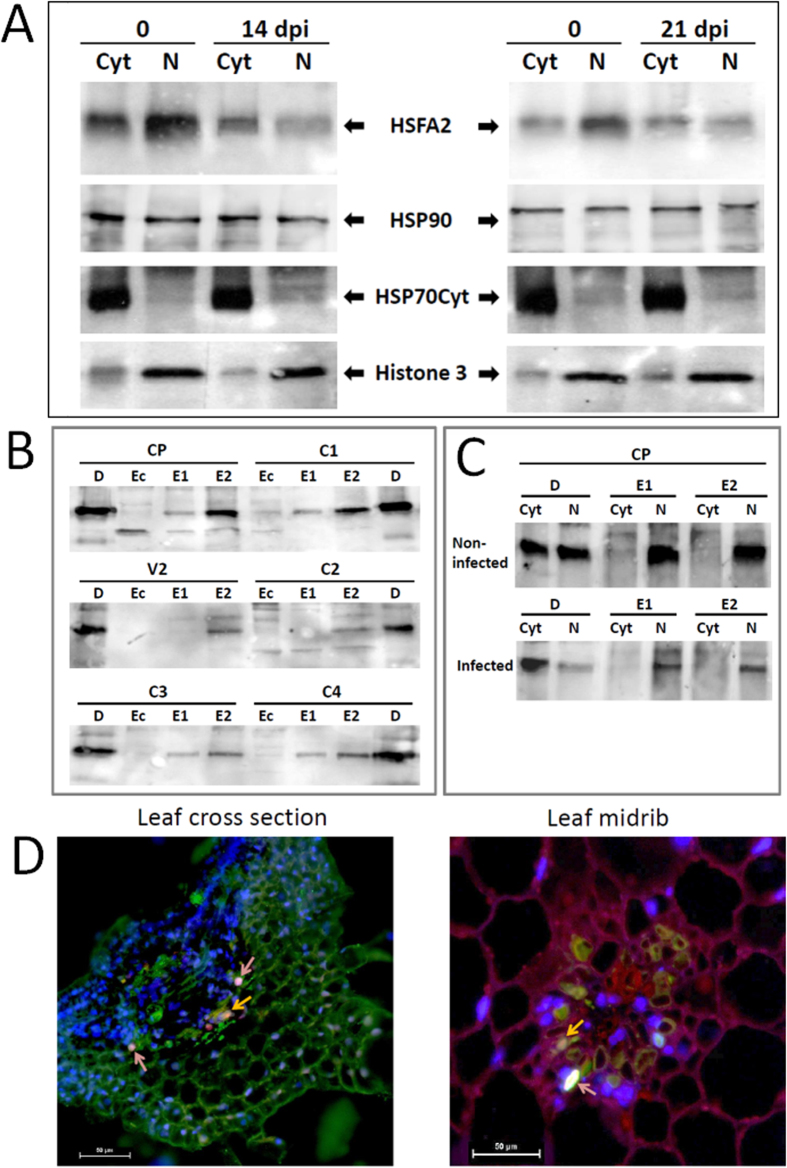
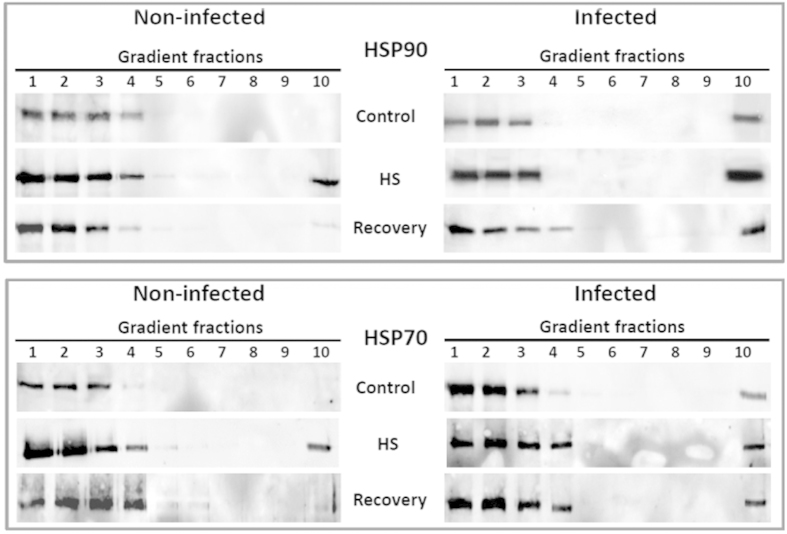
Cultured tomatoes are often exposed to a combination of extreme heat and infection with Tomato yellow leaf curl virus (TYLCV). This stress combination leads to intense disease symptoms and yield losses. The response of TYLCV-susceptible and resistant tomatoes to heat stress together with viral infection was compared. The plant heat-stress response was undermined in TYLCV infected plants. The decline correlated with the down-regulation of heat shock transcription factors (HSFs) HSFA2 and HSFB1, and consequently, of HSF-regulated genes Hsp17, Apx1, Apx2 and Hsp90. We proposed that the weakened heat stress response was due to the decreased capacity of HSFA2 to translocate into the nuclei of infected cells. All the six TYLCV proteins were able to interact with tomato HSFA2 in vitro, moreover, coat protein developed complexes with HSFA2 in nuclei. Capturing of HSFA2 by viral proteins could suppress the transcriptional activation of heat stress response genes. Application of both heat and TYLCV stresses was accompanied by the development of intracellular large protein aggregates containing TYLCV proteins and DNA. The maintenance of cellular chaperones in the aggregated state, even after recovery from heat stress, prevents the circulation of free soluble chaperones, causing an additional decrease in stress response efficiency.
Figures
References
-
- Czosnek H. Tomato Yellow Leaf Curl Virus Disease. In: Czosnek H. editor. Management, molecular biology, breeding for resistance. Dordrecht: Springer, 440p (2007).
-
- Hanley-Bowdoin L., Bejarano E. R., Robertson D. & Mansoor D. Geminiviruses: masters at redirecting and reprogramming plant processes., Nat. Rev. Microbiol. 11, 777–788 (2013). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
