

Norbin Stimulates the Catalytic Activity and Plasma Membrane Localization of the Guanine-Nucleotide Exchange Factor P-Rex1
- PMID: 26792863
- PMCID: PMC4813545
- DOI: 10.1074/jbc.M115.686592
Norbin Stimulates the Catalytic Activity and Plasma Membrane Localization of the Guanine-Nucleotide Exchange Factor P-Rex1
Abstract
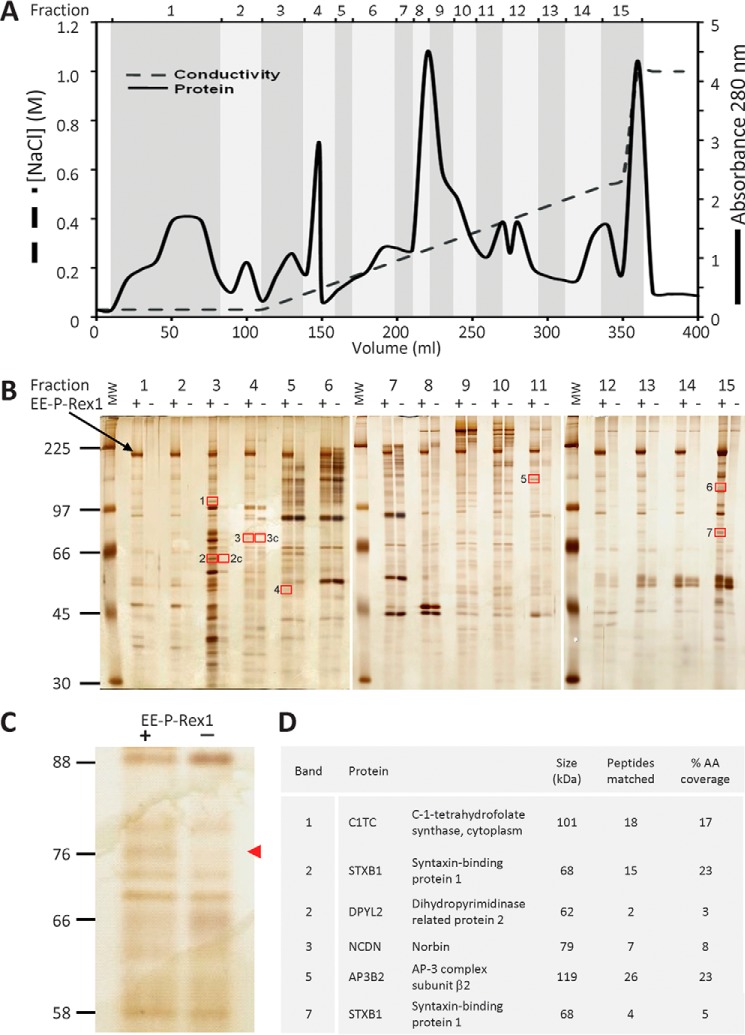
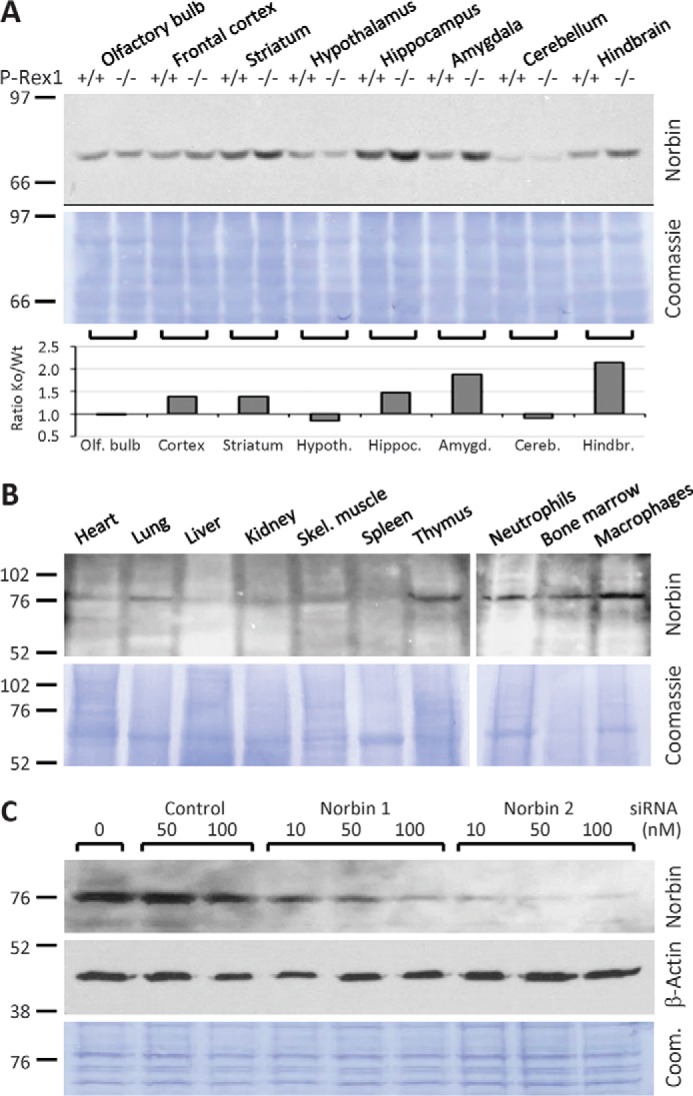
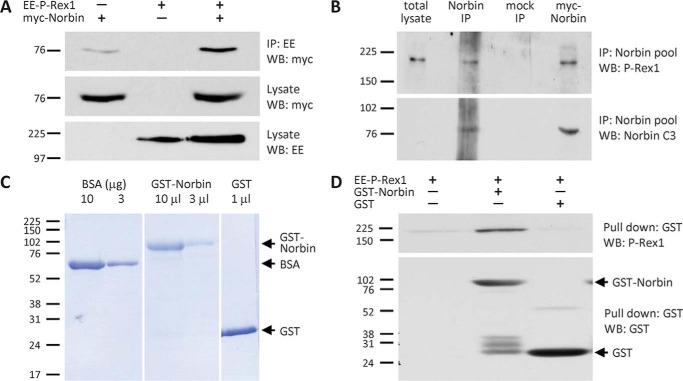
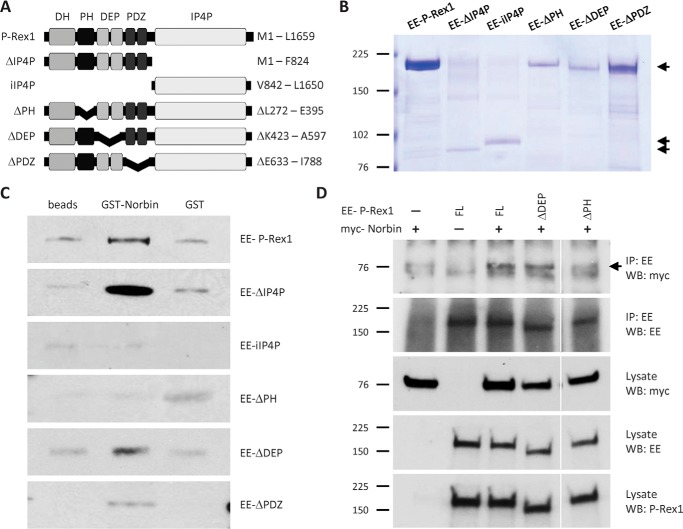
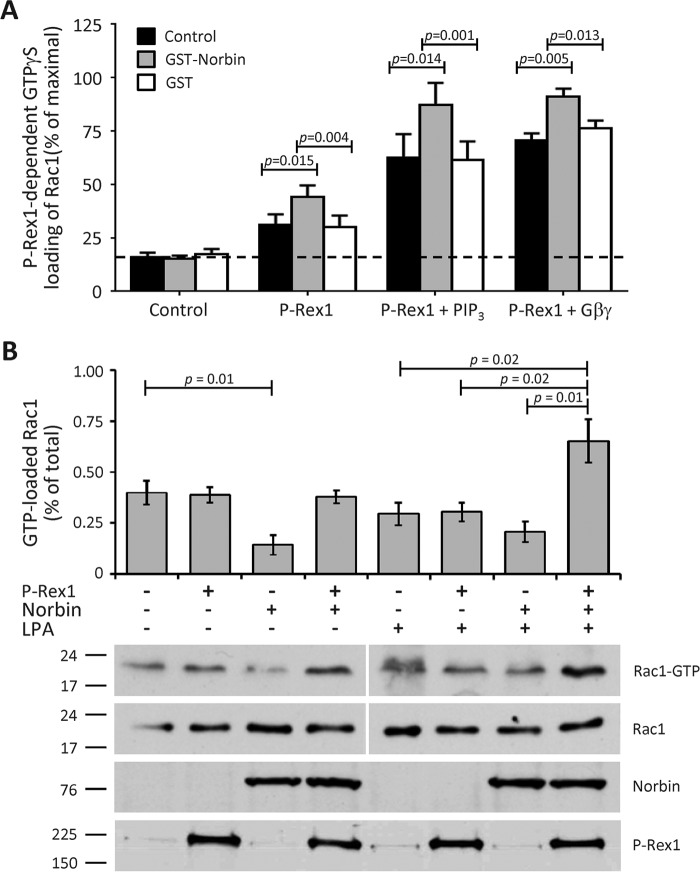
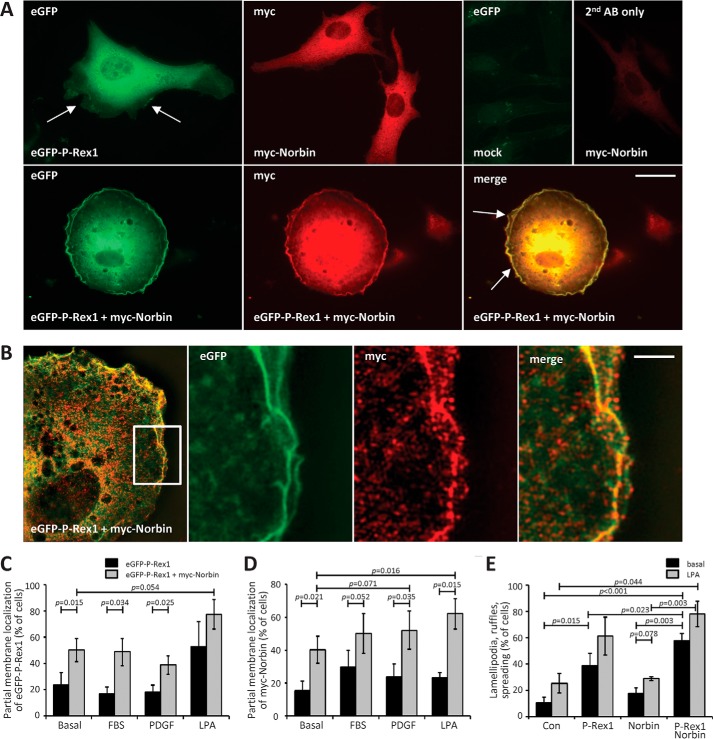
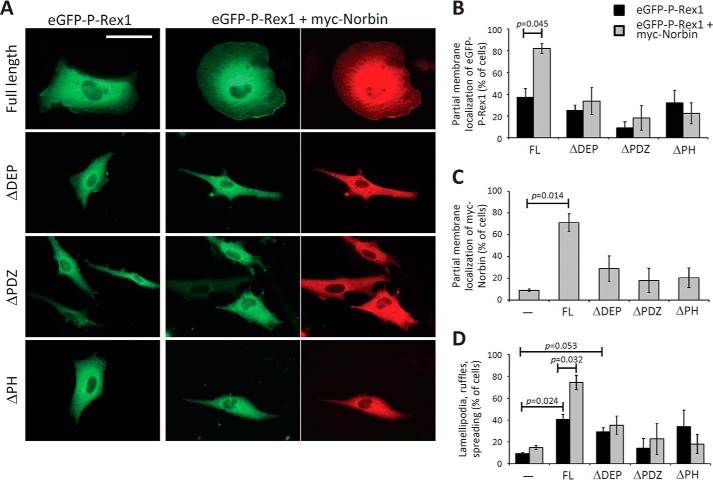
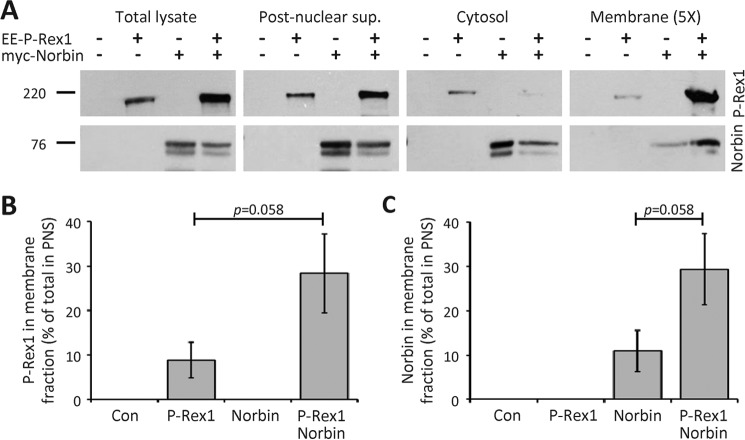
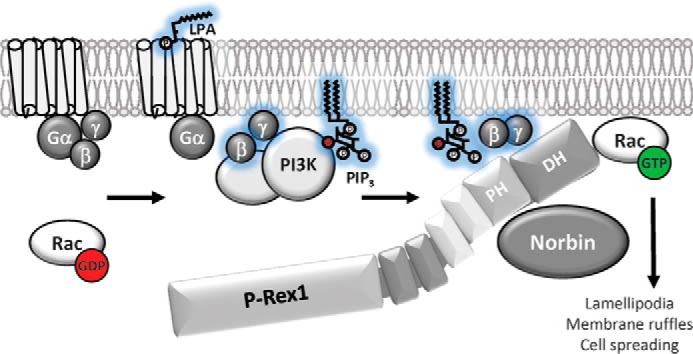
P-Rex1 is a guanine-nucleotide exchange factor (GEF) that activates the small G protein (GTPase) Rac1 to control Rac1-dependent cytoskeletal dynamics, and thus cell morphology. Three mechanisms of P-Rex1 regulation are currently known: (i) binding of the phosphoinositide second messenger PIP3, (ii) binding of the Gβγ subunits of heterotrimeric G proteins, and (iii) phosphorylation of various serine residues. Using recombinant P-Rex1 protein to search for new binding partners, we isolated the G-protein-coupled receptor (GPCR)-adaptor protein Norbin (Neurochondrin, NCDN) from mouse brain fractions. Coimmunoprecipitation confirmed the interaction between overexpressed P-Rex1 and Norbin in COS-7 cells, as well as between endogenous P-Rex1 and Norbin in HEK-293 cells. Binding assays with purified recombinant proteins showed that their interaction is direct, and mutational analysis revealed that the pleckstrin homology domain of P-Rex1 is required. Rac-GEF activity assays with purified recombinant proteins showed that direct interaction with Norbin increases the basal, PIP3- and Gβγ-stimulated Rac-GEF activity of P-Rex1. Pak-CRIB pulldown assays demonstrated that Norbin promotes the P-Rex1-mediated activation of endogenous Rac1 upon stimulation of HEK-293 cells with lysophosphatidic acid. Finally, immunofluorescence microscopy and subcellular fractionation showed that coexpression of P-Rex1 and Norbin induces a robust translocation of both proteins from the cytosol to the plasma membrane, as well as promoting cell spreading, lamellipodia formation, and membrane ruffling, cell morphologies generated by active Rac1. In summary, we have identified a novel mechanism of P-Rex1 regulation through the GPCR-adaptor protein Norbin, a direct P-Rex1 interacting protein that promotes the Rac-GEF activity and membrane localization of P-Rex1.
Keywords: G protein-coupled receptor (GPCR); Neurochondrin (NCDN); PREX1; PREX2; Rac (Rac GTPase); Rho (Rho GTPase); cell signaling; guanine nucleotide exchange factor (GEF); protein complex; small GTPase.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Welch H. C., Coadwell W. J., Ellson C. D., Ferguson G. J., Andrews S. R., Erdjument-Bromage H., Tempst P., Hawkins P. T., and Stephens L. R. (2002) P-Rex1, a PtdIns(3,4,5)P3- and Gβγ-regulated guanine-nucleotide exchange factor for Rac. Cell 108, 809–821 - PubMed
-
- Damoulakis G., Gambardella L., Rossman K. L., Lawson C. D., Anderson K. E., Fukui Y., Welch H. C., Der C. J., Stephens L. R., and Hawkins P. T. (2014) P-Rex1 directly activates RhoG to regulate GPCR-driven Rac signalling and actin polarity in neutrophils. J. Cell Sci. 127, 2589–2600 - PubMed
-
- Lawson C. D., Donald S., Anderson K. E., Patton D. T., and Welch H. (2011) P-Rex1 and Vav1 cooperate in the regulation of fMLF-dependent neutrophil responses. J. Immunol. 186, 1467–1476 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
