

Lipid Flippases for Bacterial Peptidoglycan Biosynthesis
- PMID: 26792999
- PMCID: PMC4714577
- DOI: 10.4137/LPI.S31783
Lipid Flippases for Bacterial Peptidoglycan Biosynthesis
Abstract
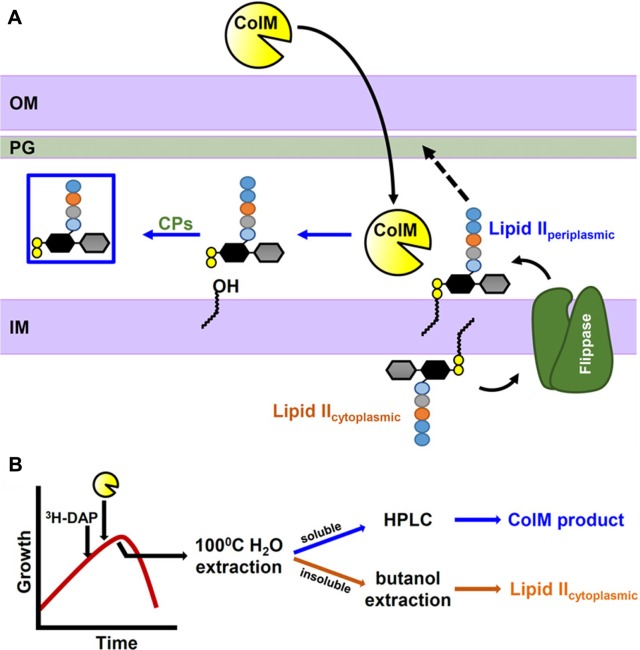
The biosynthesis of cellular polysaccharides and glycoconjugates often involves lipid-linked intermediates that need to be translocated across membranes. Essential pathways such as N-glycosylation in eukaryotes and biogenesis of the peptidoglycan (PG) cell wall in bacteria share a common strategy where nucleotide-sugars are used to build a membrane-bound oligosaccharide precursor that is linked to a phosphorylated isoprenoid lipid. Once made, these lipid-linked intermediates must be translocated across a membrane so that they can serve as substrates in a different cellular compartment. How translocation occurs is poorly understood, although it clearly requires a transporter or flippase. Identification of these transporters is notoriously difficult, and, in particular, the identity of the flippase of lipid II, an intermediate required for PG biogenesis, has been the subject of much debate. Here, I will review the body of work that has recently fueled this controversy, centered on proposed flippase candidates FtsW, MurJ, and AmJ.
Keywords: MATE transporter; MOP exporter; MviN; YdaH; murein.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
