

Thalamic Circuit Diversity: Modulation of the Driver/Modulator Framework
- PMID: 26793068
- PMCID: PMC4709853
- DOI: 10.3389/fncir.2015.00086
Thalamic Circuit Diversity: Modulation of the Driver/Modulator Framework
Abstract
The idea that dorsal thalamic inputs can be divided into "drivers", which provide the primary excitatory drive for the relay of information to cortex, and "modulators", which alter the gain of signal transmission, has provided a valuable organizing principle for the study of thalamic function. This view further promoted the identification of "first order" and "higher order" thalamic nuclei, based on the origin of their driving inputs. Since the introduction of this influential terminology, a number of studies have revealed the existence of a wide variety of thalamic organizational schemes. For example, some thalamic nuclei are not innervated by typical driver inputs, but instead receive input from terminals which exhibit features distinct from those of either classic drivers or modulators. In addition, many thalamic nuclei contain unique combinations of convergent first order, higher order, and/or other "driver-like" inputs that do not conform with the driver/modulator framework. The assortment of synaptic arrangements identified in the thalamus are reviewed and discussed from the perspective that this organizational diversity can dramatically increase the computational capabilities of the thalamus, reflecting its essential roles in sensory, motor, and sensory-motor circuits.
Keywords: corticothalamic; dorsal lateral geniculate nucleus; lateral posterior nucleus; pulvinar nucleus; retinogeniculate; tectothalamic; thalamocortical.
Figures
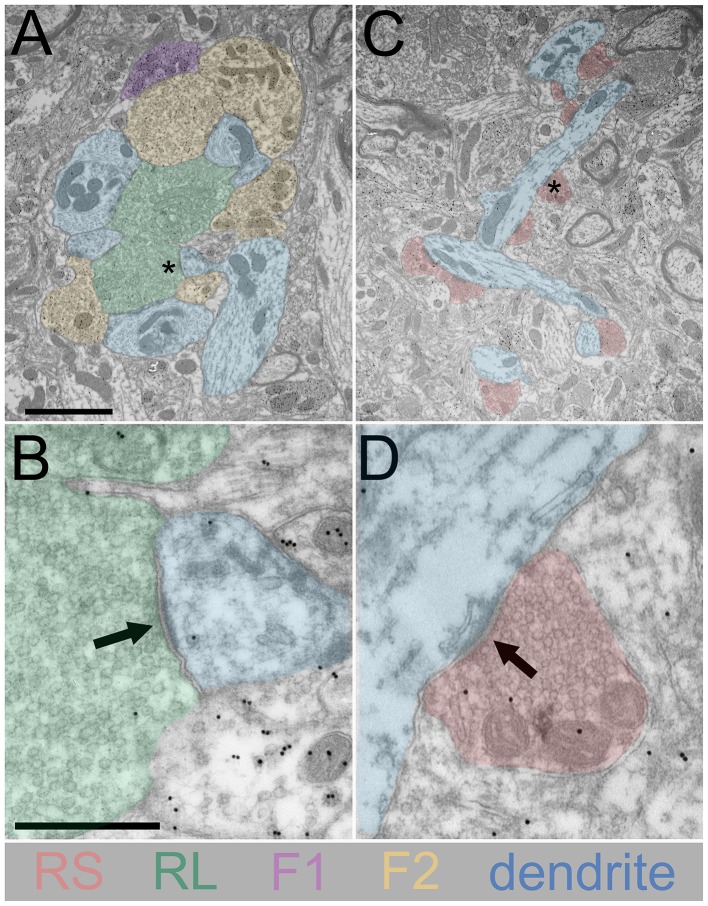
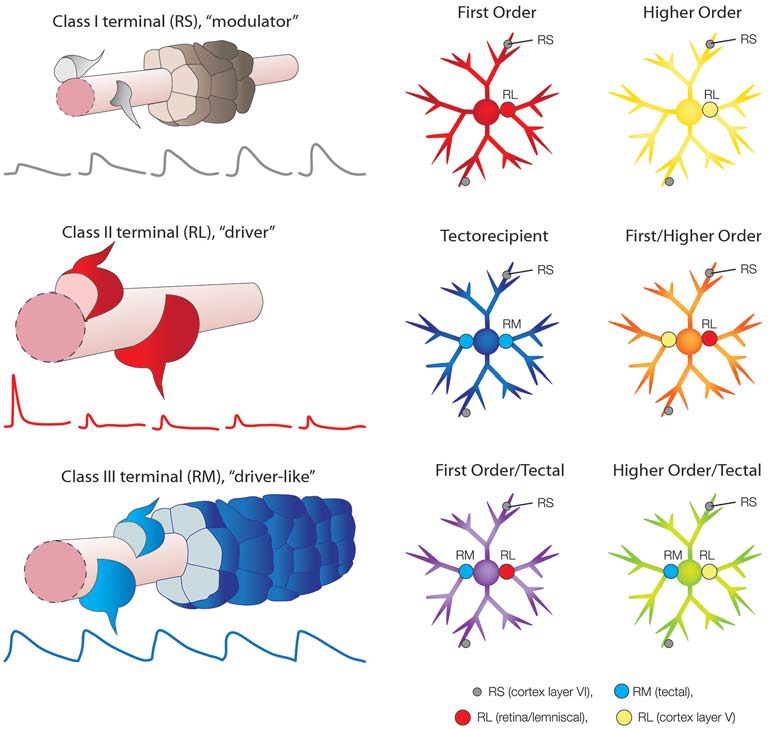
Similar articles
-
Thalamic relays and cortical functioning.Prog Brain Res. 2005;149:107-26. doi: 10.1016/S0079-6123(05)49009-3. Prog Brain Res. 2005. PMID: 16226580 Review.
-
The role of the thalamus in the flow of information to the cortex.Philos Trans R Soc Lond B Biol Sci. 2002 Dec 29;357(1428):1695-708. doi: 10.1098/rstb.2002.1161. Philos Trans R Soc Lond B Biol Sci. 2002. PMID: 12626004 Free PMC article. Review.
-
Drivers from the deep: the contribution of collicular input to thalamocortical processing.Prog Brain Res. 2005;149:207-25. doi: 10.1016/S0079-6123(05)49015-9. Prog Brain Res. 2005. PMID: 16226586 Review.
-
Functioning of Circuits Connecting Thalamus and Cortex.Compr Physiol. 2017 Mar 16;7(2):713-739. doi: 10.1002/cphy.c160032. Compr Physiol. 2017. PMID: 28333385 Review.
-
Somatosensory corticothalamic projections: distinguishing drivers from modulators.J Neurophysiol. 2004 Oct;92(4):2185-97. doi: 10.1152/jn.00322.2004. Epub 2004 May 12. J Neurophysiol. 2004. PMID: 15140908
Cited by
-
Corticothalamic Pathways From Layer 5: Emerging Roles in Computation and Pathology.Front Neural Circuits. 2021 Sep 9;15:730211. doi: 10.3389/fncir.2021.730211. eCollection 2021. Front Neural Circuits. 2021. PMID: 34566583 Free PMC article. Review.
-
Not a one-trick pony: Diverse connectivity and functions of the rodent lateral geniculate complex.Vis Neurosci. 2017 Jan;34:E012. doi: 10.1017/S0952523817000098. Vis Neurosci. 2017. PMID: 28965517 Free PMC article. Review.
-
Immunocytochemical and ultrastructural organization of the taste thalamus of the tree shrew (Tupaia belangeri).J Comp Neurol. 2021 Jul 1;529(10):2558-2575. doi: 10.1002/cne.25109. Epub 2021 Jan 28. J Comp Neurol. 2021. PMID: 33458823 Free PMC article.
-
Integrated Classification of Cortical Cells and Quantitative Projectomic Mapping Unveil Organizational Principles of Brain-Wide Connectomes at Single Cell Level.bioRxiv [Preprint]. 2025 May 8:2025.05.08.652699. doi: 10.1101/2025.05.08.652699. bioRxiv. 2025. PMID: 40654623 Free PMC article. Preprint.
-
Retinorecipient areas in the common marmoset (Callithrix jacchus): An image-forming and non-image forming circuitry.Front Neural Circuits. 2023 Feb 2;17:1088686. doi: 10.3389/fncir.2023.1088686. eCollection 2023. Front Neural Circuits. 2023. PMID: 36817647 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources