

A Novel Topology of Proline-rich Transmembrane Protein 2 (PRRT2): HINTS FOR AN INTRACELLULAR FUNCTION AT THE SYNAPSE
- PMID: 26797119
- PMCID: PMC4813553
- DOI: 10.1074/jbc.M115.683888
A Novel Topology of Proline-rich Transmembrane Protein 2 (PRRT2): HINTS FOR AN INTRACELLULAR FUNCTION AT THE SYNAPSE
Erratum in
-
A novel topology of proline-rich transmembrane protein 2 (PRRT2): Hints for an intracellular function at the synapse.J Biol Chem. 2018 Mar 23;293(12):4581. doi: 10.1074/jbc.AAC118.002651. J Biol Chem. 2018. PMID: 29572331 Free PMC article. No abstract available.
Abstract
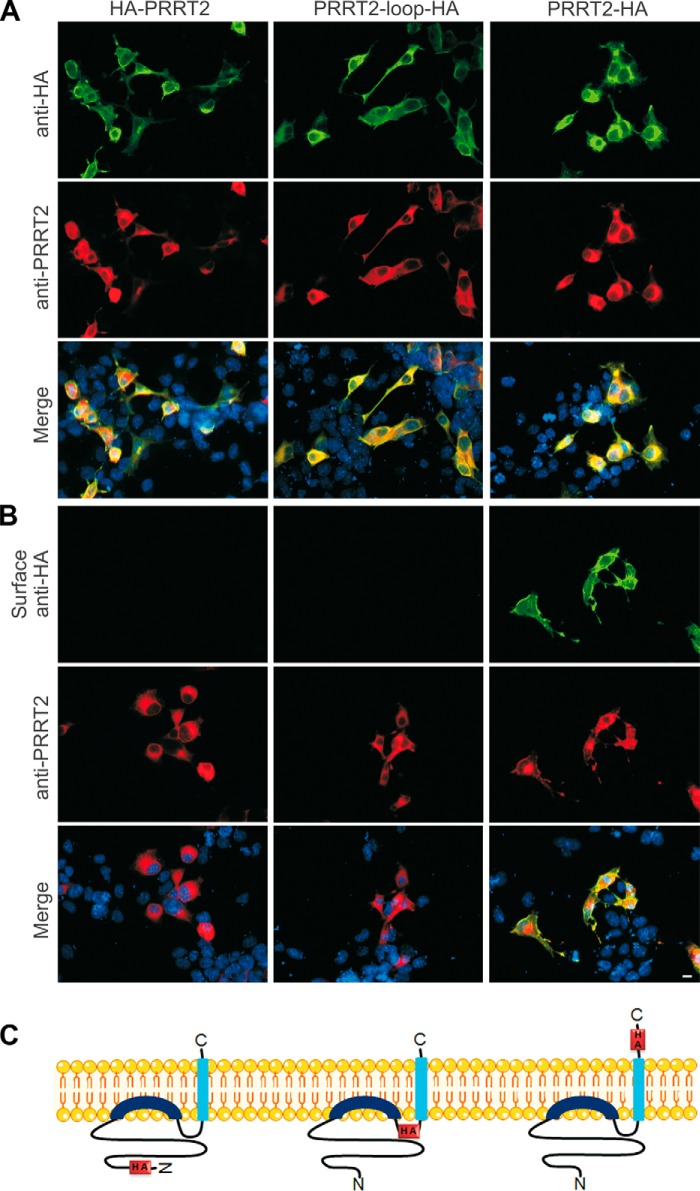
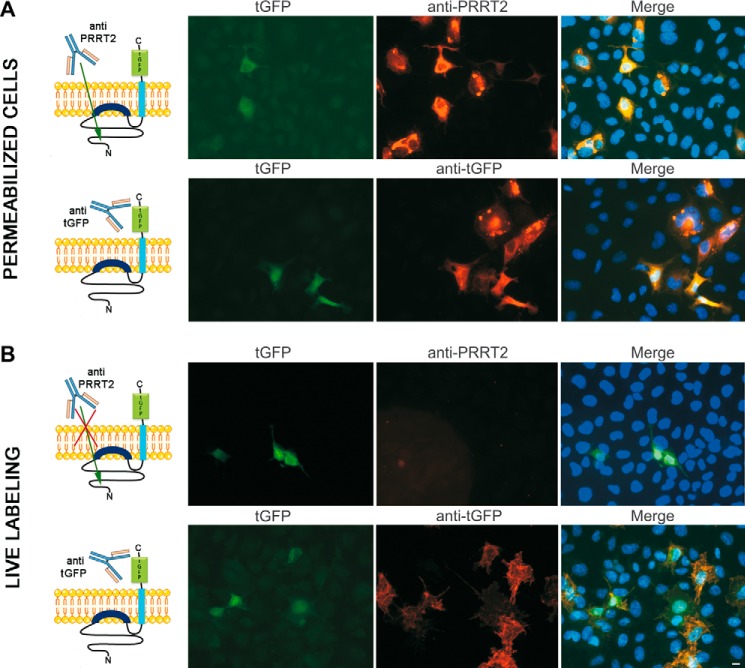
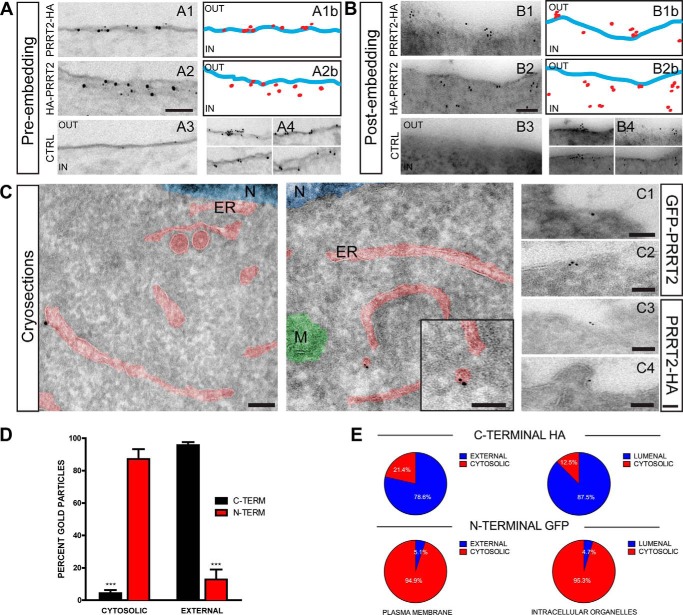
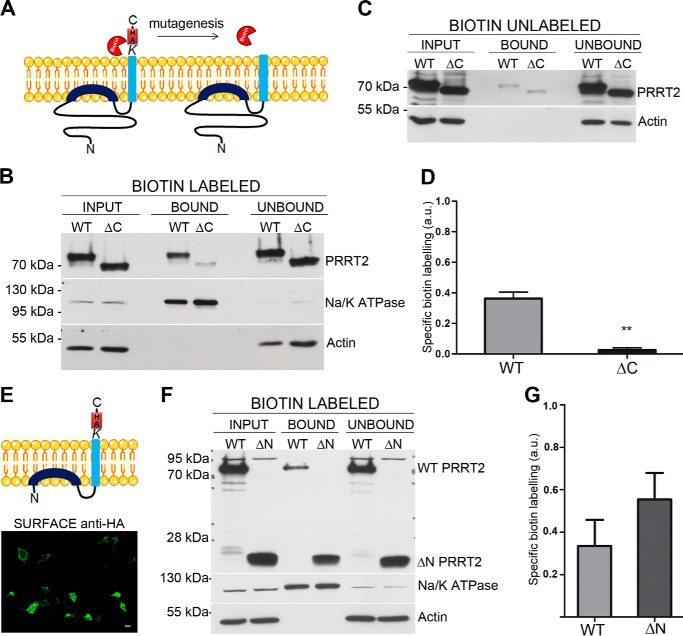
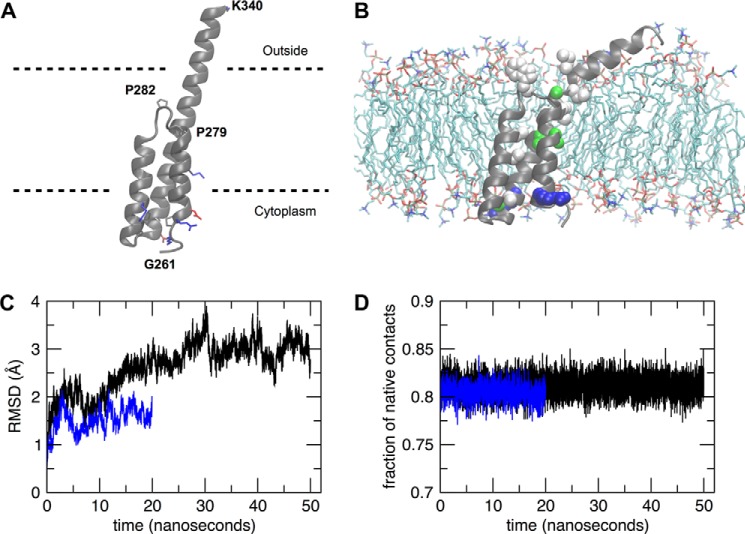
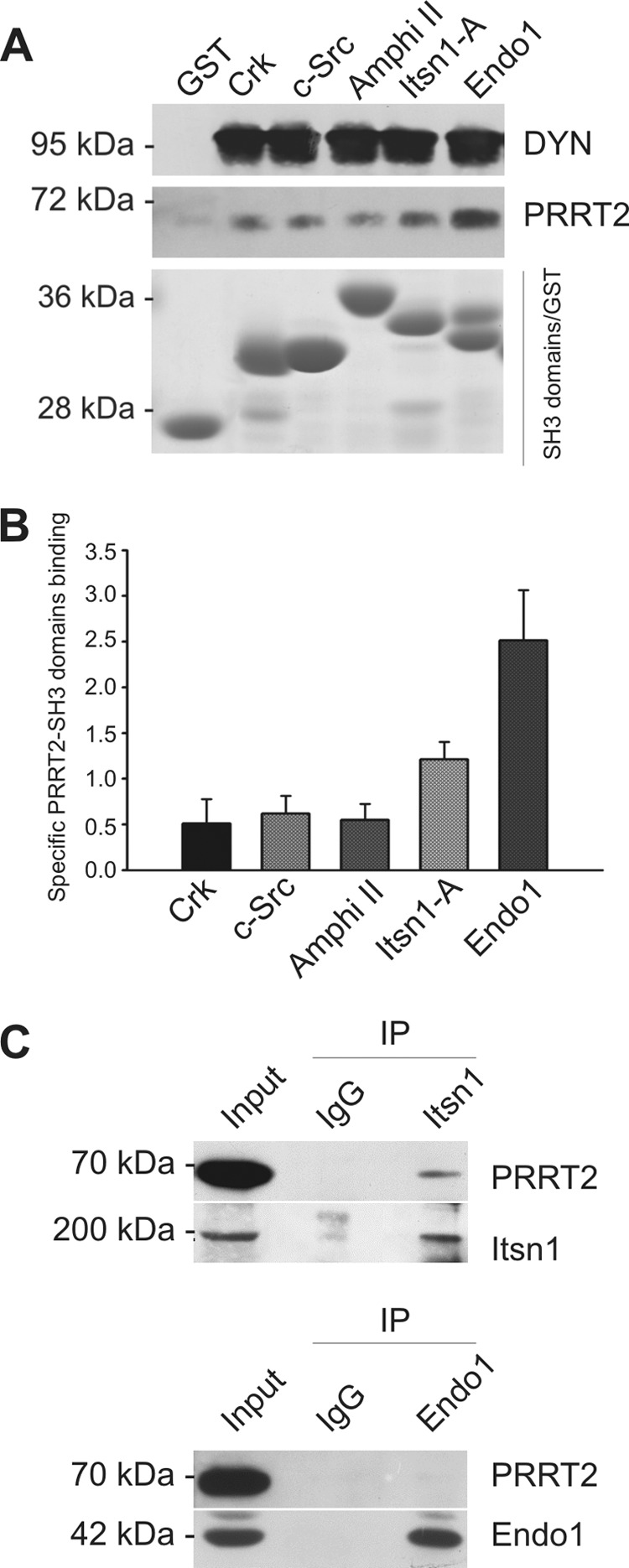
Proline-rich transmembrane protein 2 (PRRT2) has been identified as the single causative gene for a group of paroxysmal syndromes of infancy, including epilepsy, paroxysmal movement disorders, and migraine. On the basis of topology predictions, PRRT2 has been assigned to the recently characterized family of Dispanins, whose members share the two-transmembrane domain topology with a large N terminus and short C terminus oriented toward the outside of the cell. Because PRRT2 plays a role at the synapse, it is important to confirm the exact orientation of its N and C termini with respect to the plasma membrane to get clues regarding its possible function. Using a combination of different experimental approaches, including live immunolabeling, immunogold electron microscopy, surface biotinylation and computational modeling, we demonstrate a novel topology for this protein. PRRT2 is a type II transmembrane protein in which only the second hydrophobic segment spans the plasma membrane, whereas the first one is associated with the internal surface of the membrane and forms a helix-loop-helix structure without crossing it. Most importantly, the large proline-rich N-terminal domain is not exposed to the extracellular space but is localized intracellularly, and only the short C terminus is extracellular (N cyt/C exo topology). Accordingly, we show that PRRT2 interacts with the Src homology 3 domain-bearing protein Intersectin 1, an intracellular protein involved in synaptic vesicle cycling. These findings will contribute to the clarification of the role of PRRT2 at the synapse and the understanding of pathogenic mechanisms on the basis of PRRT2-related neurological disorders.
Keywords: SNARE proteins; membrane protein; molecular dynamics; neurological disease; synapse.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Méneret A., Gaudebout C., Riant F., Vidailhet M., Depienne C., and Roze E. (2013) PRRT2 mutations and paroxysmal disorders. Eur. J. Neurol. 20, 872–878 - PubMed
-
- Heron S. E., and Dibbens L. M. (2013) Role of PRRT2 in common paroxysmal neurological disorders: a gene with remarkable pleiotropy. J. Med. Genet. 50, 133–139 - PubMed
-
- Becker F., Schubert J., Striano P., Anttonen A. K., Liukkonen E., Gaily E., Gerloff C., Müller S., Heußinger N., Kellinghaus C., Robbiano A., Polvi A., Zittel S., von Oertzen T. J., Rostasy K., Schöls L., Warner T., Münchau A., Lehesjoki A. E., Zara F., Lerche H., and Weber Y. G. (2013) PRRT2-related disorders: further PKD and ICCA cases and review of the literature. J. Neurol. 260, 1234–1244 - PubMed
-
- Guerrini R., and Mink J. W. (2012) Paroxysmal disorders associated with PRRT2 mutations shake up expectations on ion channel genes. Neurology 79, 2086–2088 - PubMed
-
- Wood H. (2012) Genetics: Expanding the spectrum of neurological disorders associated with PRRT2 mutations. Nat. Rev. Neurol. 8, 657. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
