

Developmental Patterning as a Quantitative Trait: Genetic Modulation of the Hoxb6 Mutant Skeletal Phenotype
- PMID: 26800342
- PMCID: PMC4723086
- DOI: 10.1371/journal.pone.0146019
Developmental Patterning as a Quantitative Trait: Genetic Modulation of the Hoxb6 Mutant Skeletal Phenotype
Abstract
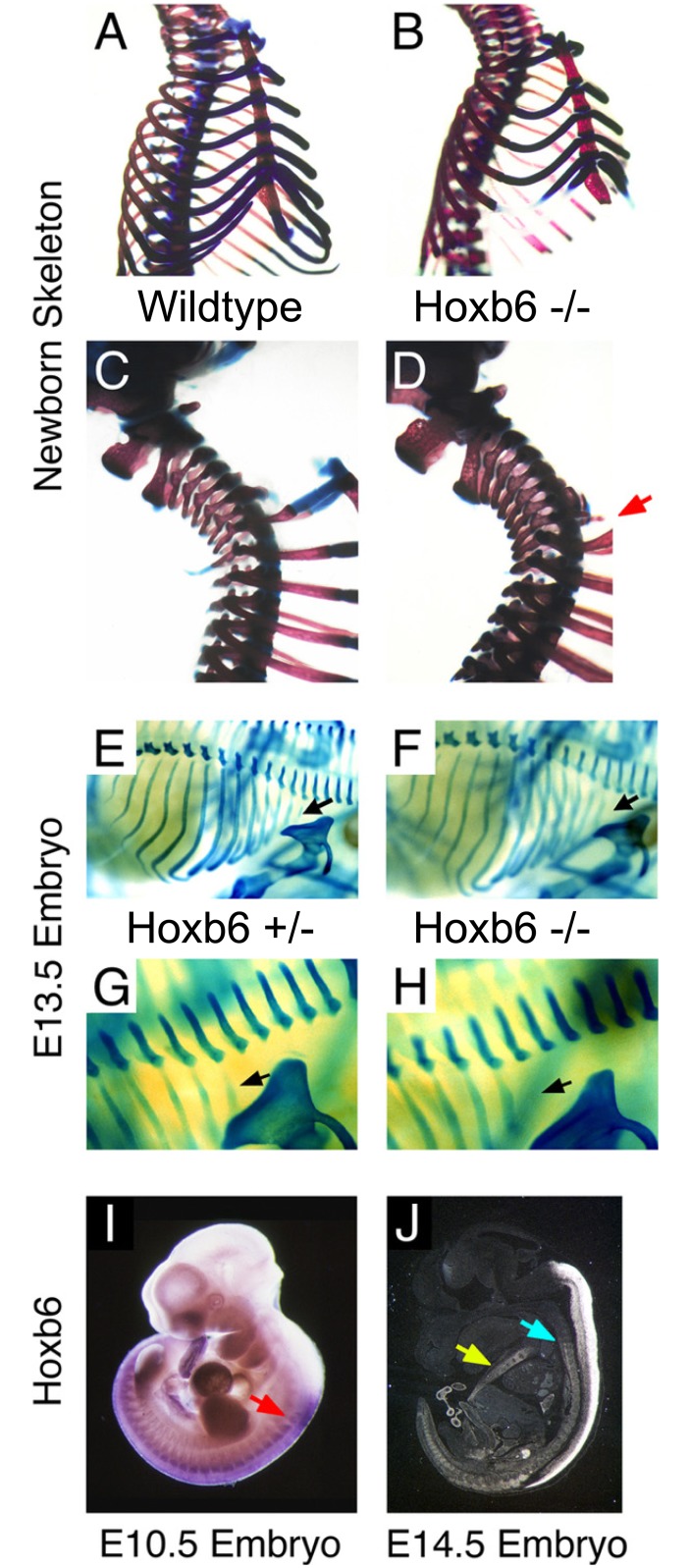
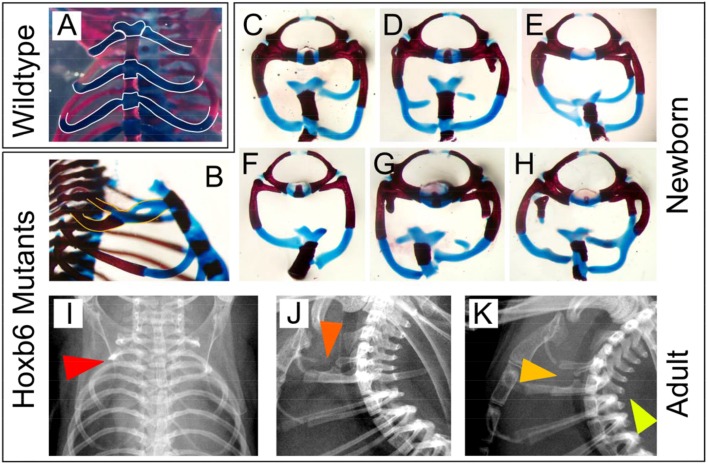
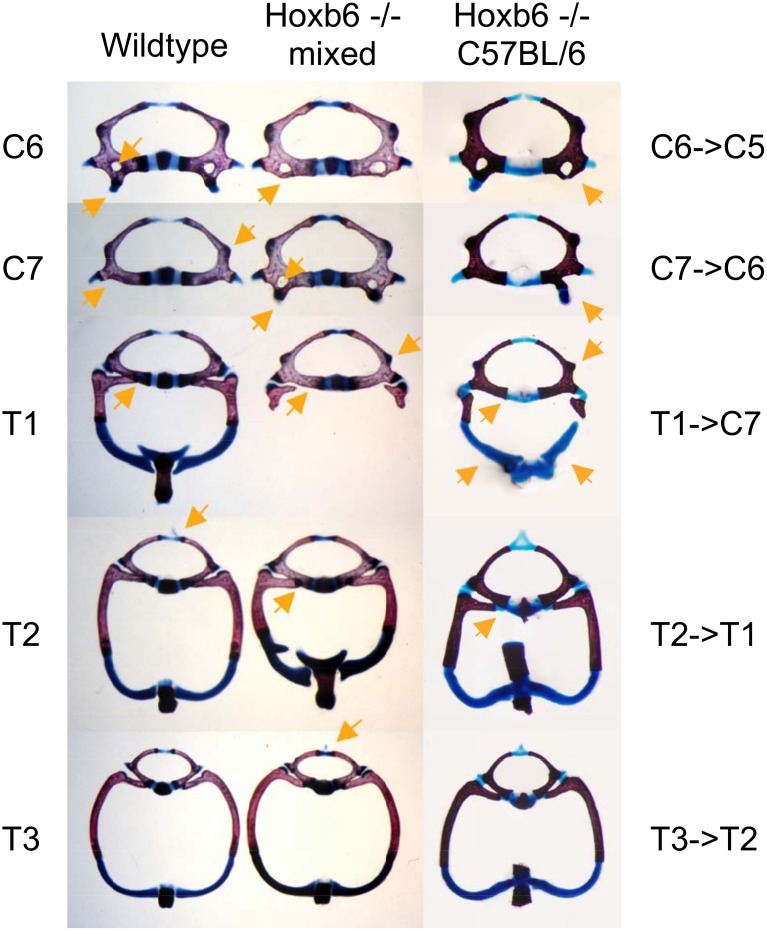
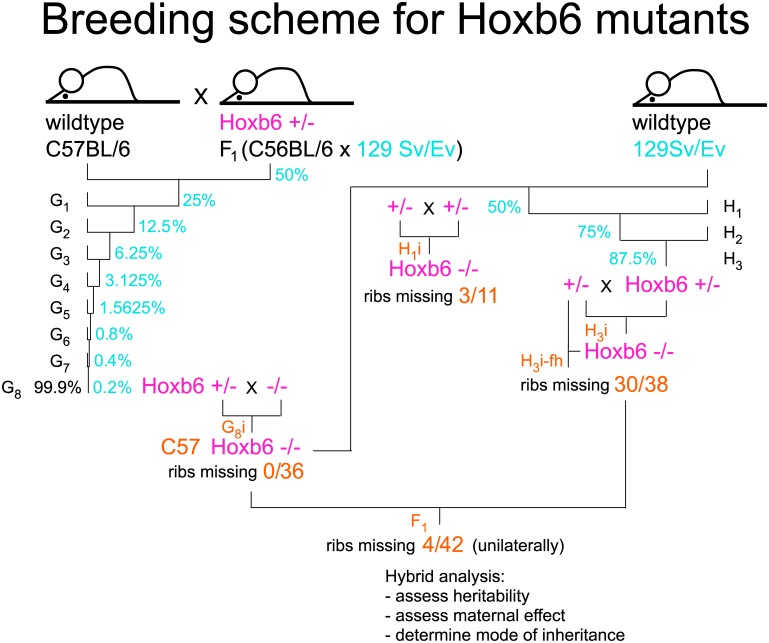
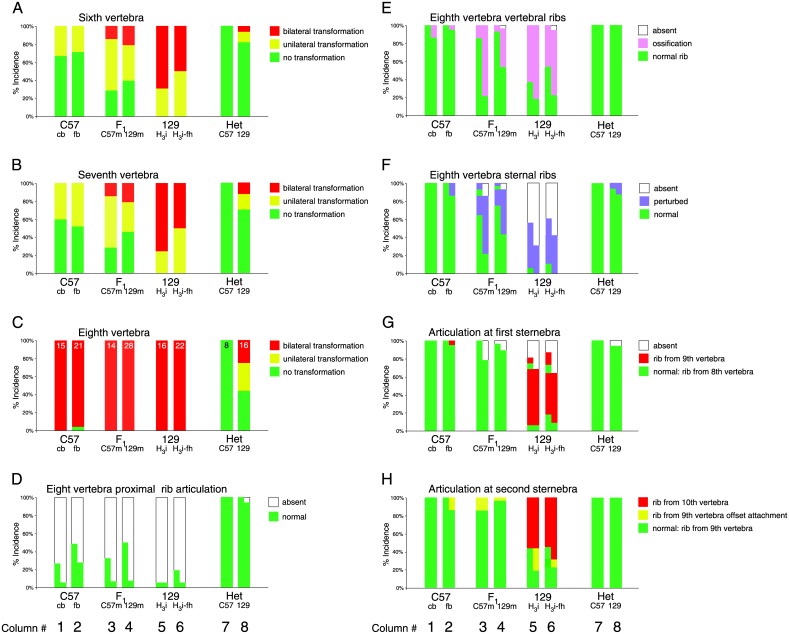
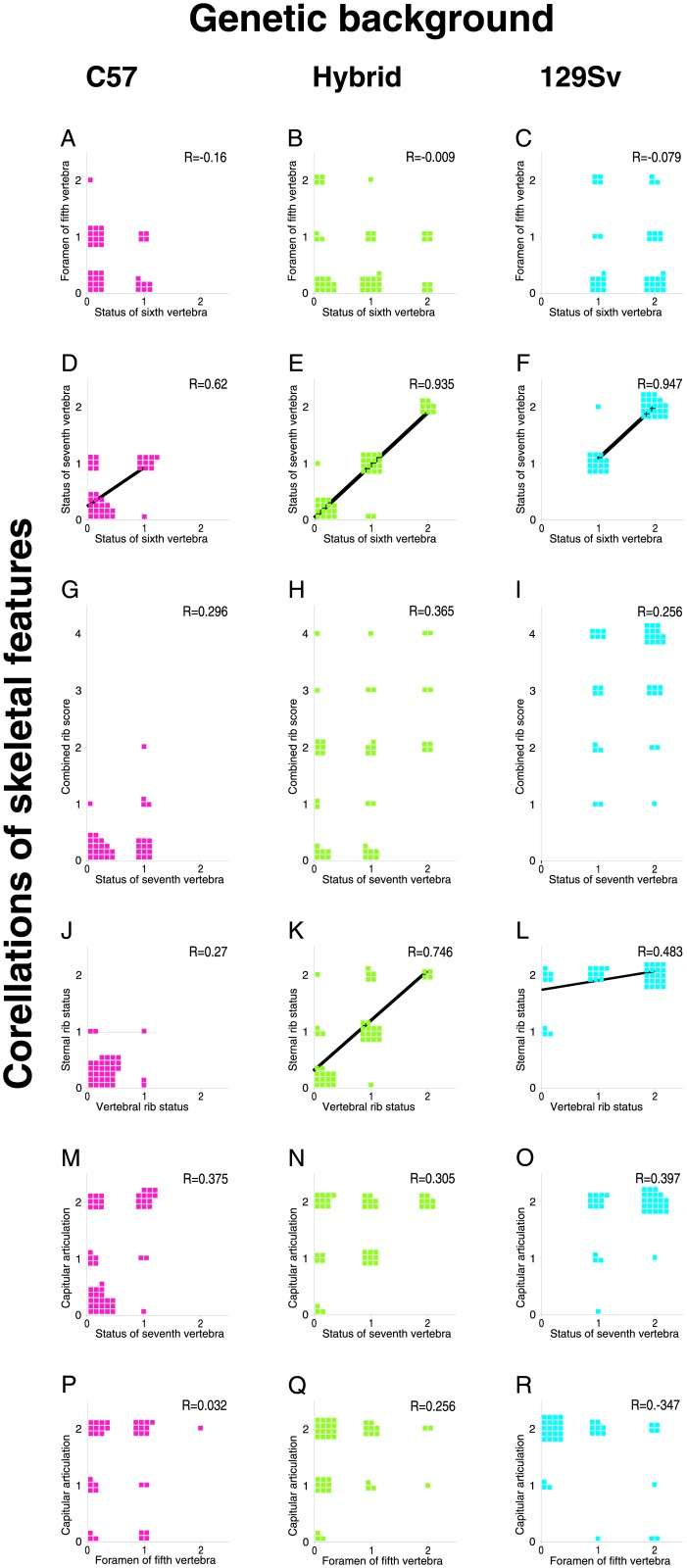
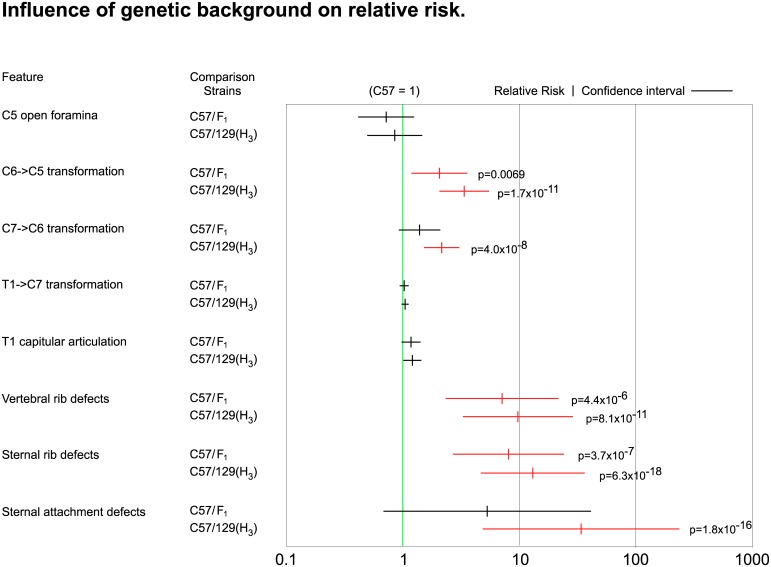
The process of patterning along the anterior-posterior axis in vertebrates is highly conserved. The function of Hox genes in the axis patterning process is particularly well documented for bone development in the vertebral column and the limbs. We here show that Hoxb6, in skeletal elements at the cervico-thoracic junction, controls multiple independent aspects of skeletal pattern, implicating discrete developmental pathways as substrates for this transcription factor. In addition, we demonstrate that Hoxb6 function is subject to modulation by genetic factors. These results establish Hox-controlled skeletal pattern as a quantitative trait modulated by gene-gene interactions, and provide evidence that distinct modifiers influence the function of conserved developmental genes in fundamental patterning processes.
Conflict of interest statement
Figures
Similar articles
-
Hox patterning of the vertebrate rib cage.Development. 2007 Aug;134(16):2981-9. doi: 10.1242/dev.007567. Epub 2007 Jul 11. Development. 2007. PMID: 17626057
-
Axial skeletal patterning in mice lacking all paralogous group 8 Hox genes.Development. 2001 May;128(10):1911-21. doi: 10.1242/dev.128.10.1911. Development. 2001. PMID: 11311170
-
Hoxb6 can interfere with somitogenesis in the posterior embryo through a mechanism independent of its rib-promoting activity.Development. 2016 Feb 1;143(3):437-48. doi: 10.1242/dev.133074. Epub 2015 Dec 30. Development. 2016. PMID: 26718008
-
Hox genes and vertebrate axial pattern.Curr Top Dev Biol. 2009;88:257-78. doi: 10.1016/S0070-2153(09)88009-5. Curr Top Dev Biol. 2009. PMID: 19651308 Review.
-
Assisting Hox proteins in controlling body form: are there new lessons from flies (and mammals)?Curr Opin Genet Dev. 2005 Aug;15(4):422-9. doi: 10.1016/j.gde.2005.06.009. Curr Opin Genet Dev. 2005. PMID: 15979870 Review.
Cited by
-
SMCHD1 has separable roles in chromatin architecture and gene silencing that could be targeted in disease.Nat Commun. 2023 Sep 25;14(1):5466. doi: 10.1038/s41467-023-40992-6. Nat Commun. 2023. PMID: 37749075 Free PMC article.
-
Effects of MLL5 and HOXA regulated by NRP1 on radioresistance in A549.Oncol Lett. 2021 May;21(5):403. doi: 10.3892/ol.2021.12664. Epub 2021 Mar 19. Oncol Lett. 2021. PMID: 33777226 Free PMC article.
-
Genomic insights into the conservation and population genetics of two Chinese native goat breeds.J Anim Sci. 2022 Oct 1;100(10):skac274. doi: 10.1093/jas/skac274. J Anim Sci. 2022. PMID: 35998083 Free PMC article.
-
Folate supplementation in three genetic models: implications for understanding folate-dependent developmental pathways.Am J Med Genet C Semin Med Genet. 2005 May 15;135C(1):24-30. doi: 10.1002/ajmg.c.30050. Am J Med Genet C Semin Med Genet. 2005. PMID: 15800896 Free PMC article. Review.
-
Examination of homozygosity runs and selection signatures in native goat breeds of Henan, China.BMC Genomics. 2024 Dec 7;25(1):1184. doi: 10.1186/s12864-024-11098-0. BMC Genomics. 2024. PMID: 39643897 Free PMC article.
References
-
- Capecchi MR. Hox genes and mammalian development. Cold Spring Harb Symp Quant Biol. 1997;62:273–281. - PubMed
-
- Davis AP, Capecchi MR. Axial homeosis and appendicular skeleton defects in mice with a targeted disruption of hoxd-11. Development. 1994;120:2187–2198. - PubMed
-
- Kostic D, Capecchi MR. Targeted disruptions of the murine hoxa-4 and hoxa-6 genes result in homeotic transformations of components of the vertebral column. Mechanisms of Development. 1994;46:231–247. - PubMed
-
- Ramirez-Solis R, Zheng H, Whiting J, Krumlauf R, Bradley A. Hoxb-4 (hox-2.6) mutant mice show homeotic transformation of a cervical vertebra and defects in the closure of the sternal rudiments. Cell. 1993;73:279–294. - PubMed
-
- Rancourt DE, Tsuzuki T, Capecchi MR. Genetic interaction between hoxb-5 and hoxb-6 is revealed by nonallelic noncomplementation. Genes Dev. 1995;9:108–122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
