

CTLA4Fcε, a novel soluble fusion protein that binds B7 molecules and the IgE receptors, and reduces human in vitro soluble CD23 production and lymphocyte proliferation
- PMID: 26801967
- PMCID: PMC4819142
- DOI: 10.1111/imm.12586
CTLA4Fcε, a novel soluble fusion protein that binds B7 molecules and the IgE receptors, and reduces human in vitro soluble CD23 production and lymphocyte proliferation
Abstract
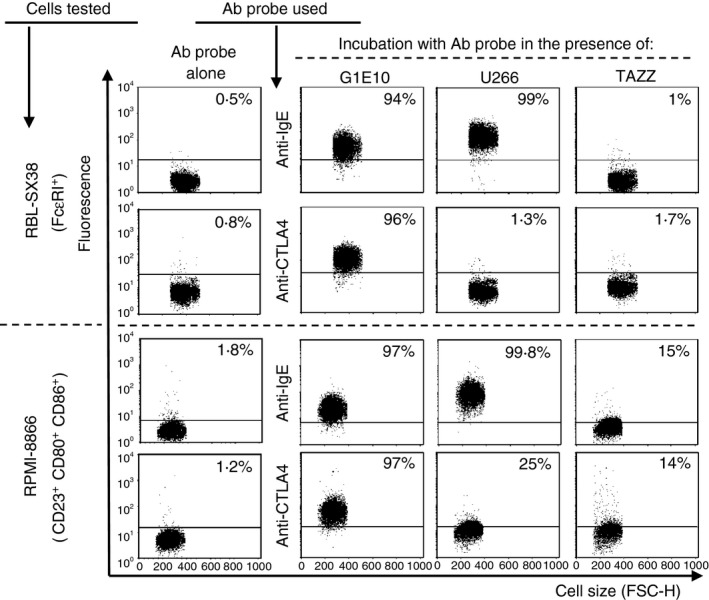
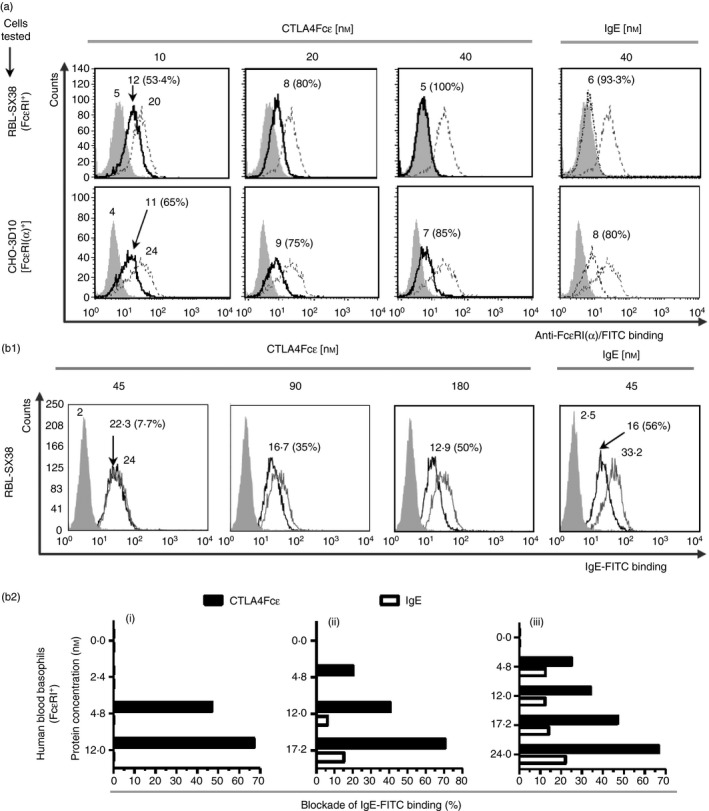
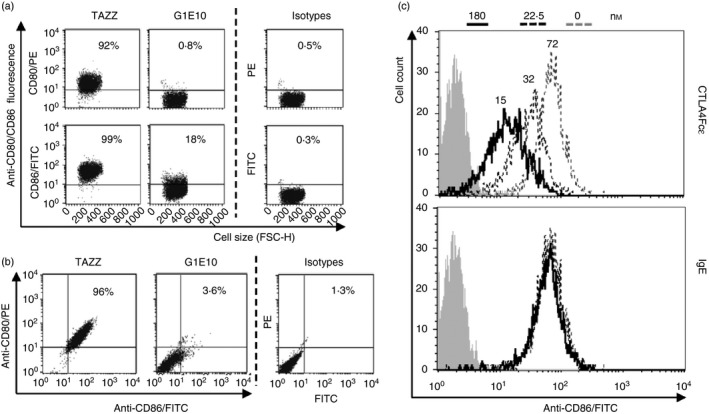
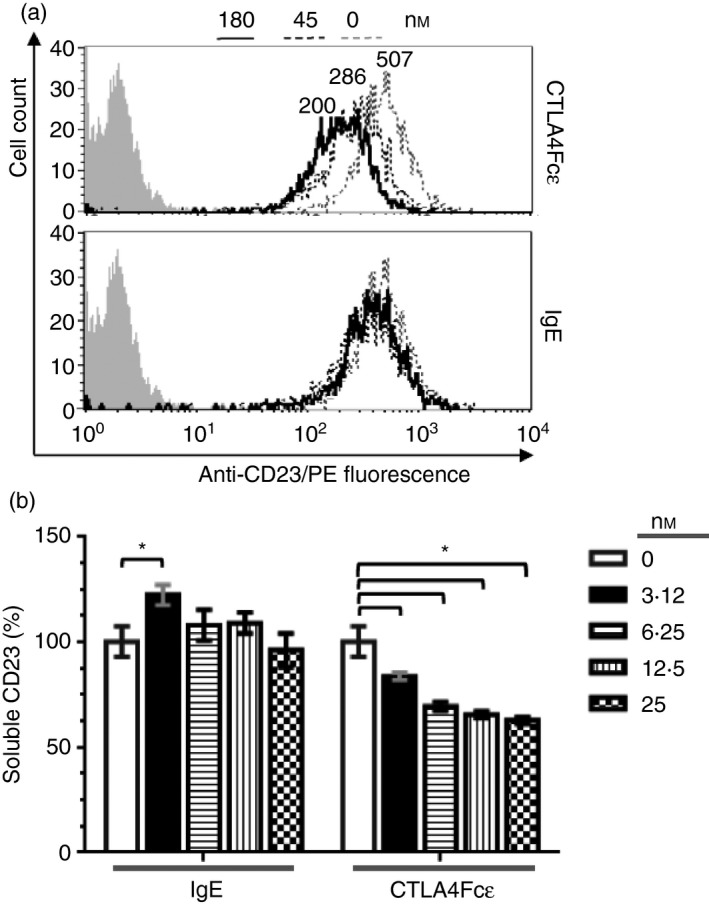
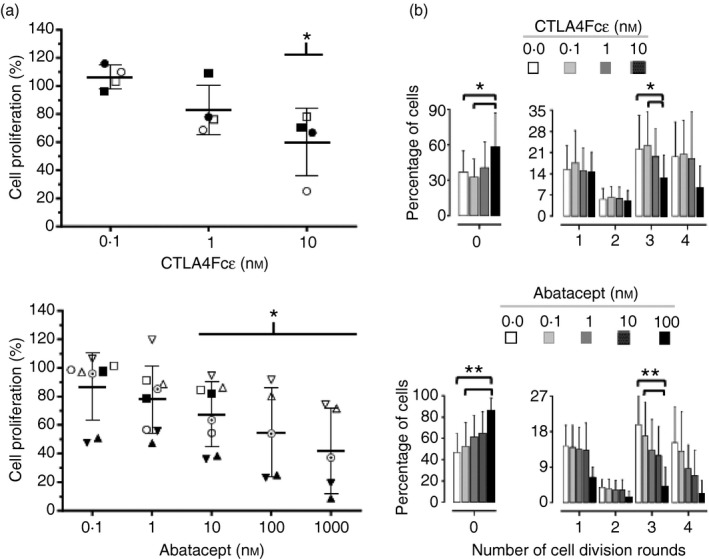
Immunoglobulin E-mediated allergy and certain autoimmune diseases are characterized by the presence of a T helper type 2 (Th2) immune response and allergen-specific or self-reactive IgE. Soluble CD23 (sCD23) is a B-cell factor that fosters IgE class-switching and synthesis, suggesting that sCD23 may be a therapeutic target for these pathologies. We produced a recombinant protein, CTLA4Fcε, by fusing the ectodomain of the immunoregulatory molecule cytotoxic T-lymphocyte antigen 4 (CTLA-4) with a fragment of the IgE H-chain constant region. In SDS-PAGE/inmunoblot analyses, CTLA4Fcε appeared as a 70,000 MW polypeptide that forms homodimers. Flow cytometry showed that CTLA4Fcε binds to IgE receptors FcεRI and FcεRII/CD23, as well as to CTLA-4 counter-receptors CD80 and CD86. Binding of CTLA4Fcε to FcεRII/CD23 appeared stronger than that of IgE. Since the cells used to study CD23 binding express CD80 and CD86, simultaneous binding of CTLA4Fcε to CD23 and CD80/CD86 seems to occur and would explain this difference. As measured by a human CD23-specific ELISA, CTLA4Fcε - but not IgE - induced a concentration-dependent reduction of sCD23 in culture supernatants of RPMI-8866 cells. Our results suggest that the simultaneous binding of CTLA4Fcɛ to CD23-CD80/CD86 may cause the formation of multi-molecular complexes that are either internalized or pose a steric hindrance to enzymatic proteolysis, so blocking sCD23 generation. CTLA4Fcε caused a concentration-dependent reduction of lymphocyte proliferation in human peripheral blood mononuclear cell samples stimulated in vitro with concanavalin A. The ability to bind IgE receptors on effector cells, to regulate the production of sCD23 and to inhibit lymphocyte proliferation suggests that CTLA4Fcɛ has immunomodulatory properties on human Th2 responses.
Keywords: CD23; CTLA-4; IgE; T helper type 2 response; fusion protein.
© 2016 John Wiley & Sons Ltd.
Figures


Similar articles
-
Soluble CD23 controls IgE synthesis and homeostasis in human B cells.J Immunol. 2012 Apr 1;188(7):3199-207. doi: 10.4049/jimmunol.1102689. Epub 2012 Mar 5. J Immunol. 2012. PMID: 22393152 Free PMC article.
-
Soluble CD23 containing B cell supernatants induce IgE from peripheral blood B-lymphocytes and costimulate with interleukin-4 in induction of IgE.J Allergy Clin Immunol. 1990 Sep;86(3 Pt 1):333-44. doi: 10.1016/s0091-6749(05)80096-x. J Allergy Clin Immunol. 1990. PMID: 1698844
-
CD86 (B7-2) on human B cells. A functional role in proliferation and selective differentiation into IgE- and IgG4-producing cells.J Biol Chem. 1997 Jun 20;272(25):15613-9. doi: 10.1074/jbc.272.25.15613. J Biol Chem. 1997. PMID: 9188449
-
The role of CD23 in the regulation of allergic responses.Allergy. 2021 Jul;76(7):1981-1989. doi: 10.1111/all.14724. Epub 2021 Jan 16. Allergy. 2021. PMID: 33378583 Free PMC article. Review.
-
CD23/FcεRII: molecular multi-tasking.Clin Exp Immunol. 2010 Oct;162(1):12-23. doi: 10.1111/j.1365-2249.2010.04210.x. Clin Exp Immunol. 2010. PMID: 20831712 Free PMC article. Review.
Cited by
-
Research Advances in Fusion Protein-Based Drugs for Diabetes Treatment.Diabetes Metab Syndr Obes. 2024 Jan 23;17:343-362. doi: 10.2147/DMSO.S421527. eCollection 2024. Diabetes Metab Syndr Obes. 2024. PMID: 38288338 Free PMC article. Review.
-
Tracing IgE-Producing Cells in Allergic Patients.Cells. 2019 Aug 28;8(9):994. doi: 10.3390/cells8090994. Cells. 2019. PMID: 31466324 Free PMC article. Review.
-
Anti-IgE therapy for IgE-mediated allergic diseases: from neutralizing IgE antibodies to eliminating IgE+ B cells.Clin Transl Allergy. 2018 Jul 18;8:27. doi: 10.1186/s13601-018-0213-z. eCollection 2018. Clin Transl Allergy. 2018. PMID: 30026908 Free PMC article. Review.
References
-
- Forsthuber TG, Ji N. Quo vadis Th1 and Th2 cells in autoimmunity and infectious diseases: Th17 cells, the new kid on the block. Exp Rev Clin Immunol 2007; 3:251–4. - PubMed
-
- Holgate S, Casale T, Wenzel S, Bousquet J, Deniz Y, Reisner C. The anti‐inflammatory effects of omalizumab confirm the central role of IgE in allergic inflammation. J Allergy Clin Immunol 2005; 115:459–65. - PubMed
-
- Nagel A, Lang A, Engel D, Podstawa E, Hunzelmann N, de Pita O et al Clinical activity of pemphigus vulgaris relates to IgE autoantibodies against desmoglein 3. Clin Immunol 2010; 134:320–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
