

Divergence and Functional Degradation of a Sex Chromosome-like Supergene
- PMID: 26804558
- PMCID: PMC4747794
- DOI: 10.1016/j.cub.2015.11.069
Divergence and Functional Degradation of a Sex Chromosome-like Supergene
Abstract
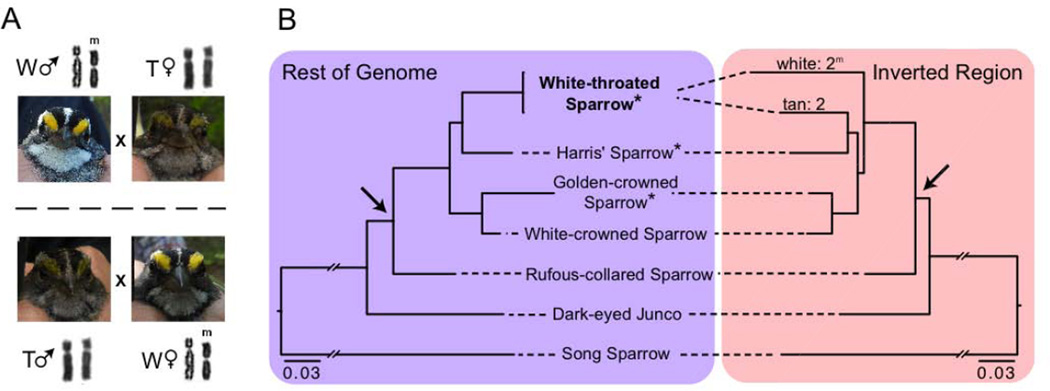
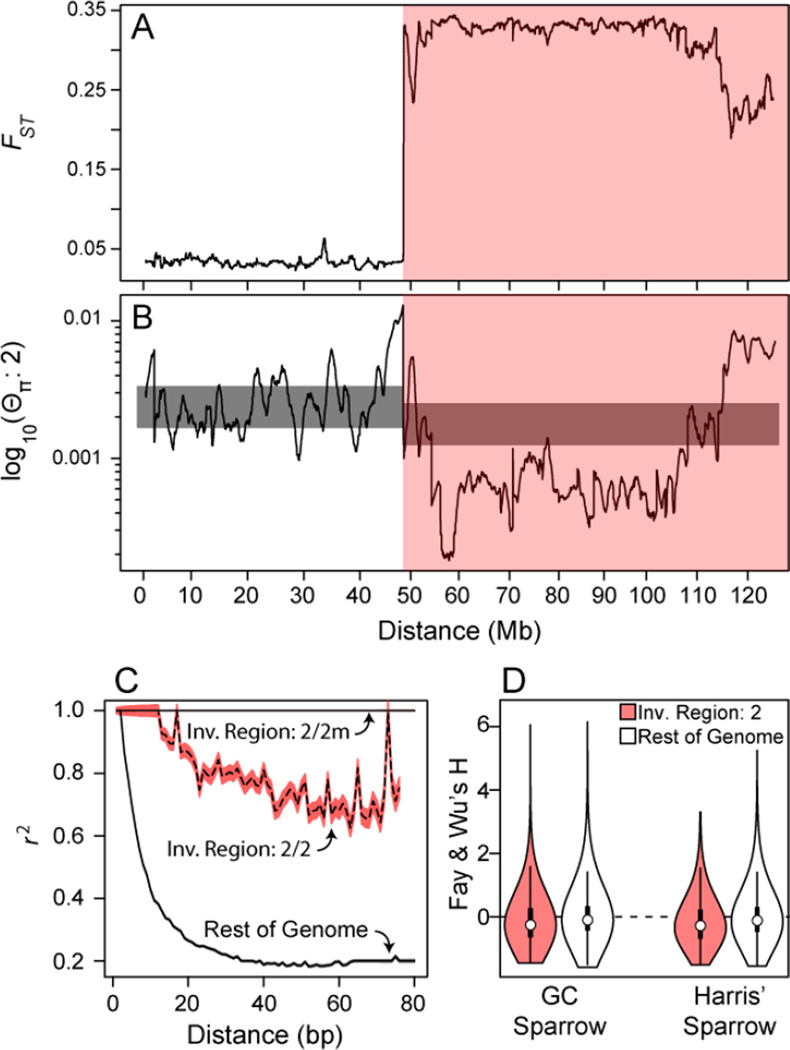
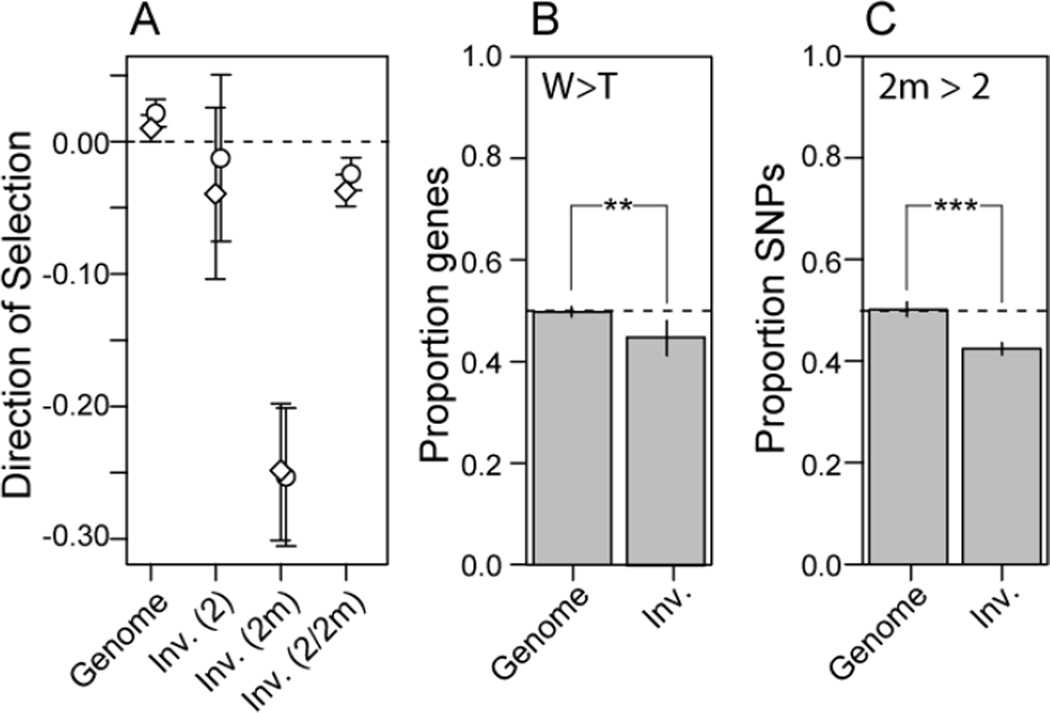
A major challenge in biology is to understand the genetic basis of adaptation. One compelling idea is that groups of tightly linked genes (i.e., "supergenes" [1, 2]) facilitate adaptation in suites of traits that determine fitness. Despite their likely importance, little is known about how alternate supergene alleles arise and become differentiated, nor their ultimate fate within species. Herein we address these questions by investigating the evolutionary history of a supergene in white-throated sparrows, Zonotrichia albicollis. This species comprises two morphs, tan and white, that differ in pigmentation and components of social behavior [3-5]. Morph is determined by alternative alleles at a balanced >100-Mb inversion-based supergene, providing a unique system for studying gene-behavior relationships. Using over two decades of field data, we document near-perfect disassortative mating among morphs, as well as the fitness consequences of rare assortative mating. We use de novo whole-genome sequencing coupled with population- and phylogenomic data to show that alternate supergene alleles are highly divergent at over 1,000 genes and that these alleles originated prior to the split of Z. albicollis from its sister species and may be polymorphic in Z. albicollis due to a past hybridization event. We provide evidence that the "white" allele may be degrading, similar to neo-Y/W sex chromosomes. We further show that the "tan" allele has surprisingly low levels of genetic diversity yet does not show several canonical signatures of recurrent positive selection. We discuss these results in the context of the origin, molecular evolution, and possible fate of this remarkable polymorphism.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Supergenes: The Genomic Architecture of a Bird with Four Sexes.Curr Biol. 2016 Feb 8;26(3):R105-7. doi: 10.1016/j.cub.2015.12.005. Curr Biol. 2016. PMID: 26859263
References
-
- Kunte K, Zhang W, Tenger-Trolander A, Palmer DH, Martin A, Reed RD, Mullen SP, Kronforst MR. Doublesex is a mimicry supergene. Nature. 2014;507:229–232. - PubMed
-
- Wang J, Wurm Y, Nipitwattanaphon M, Riba-Grognuz O, Huang Y-C, Shoemaker, Keller L. A Y-like social chromosome causes alternative colony organization in fire ants. Nature. 2013;494:664–668. - PubMed
-
- Tuttle EM. Alternative reproductive strategies in the white-throated sparrow: behavioral and genetic evidence. Behav Ecol. 2003;14:425–432.
-
- Formica VA, Tuttle EM. Examining the social landscapes of alternative reproductive strategies. J Evol Biol. 2009;22:2395–2408. - PubMed
-
- Knapton RW, Falls JB. Differences in parental contribution among pair types in the polymorphic white-throated sparrow (Zonotrichia albicollis) Can J Zool. 1983;61:1288–1292.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
