

Mutational Strand Asymmetries in Cancer Genomes Reveal Mechanisms of DNA Damage and Repair
- PMID: 26806129
- PMCID: PMC4753048
- DOI: 10.1016/j.cell.2015.12.050
Mutational Strand Asymmetries in Cancer Genomes Reveal Mechanisms of DNA Damage and Repair
Abstract
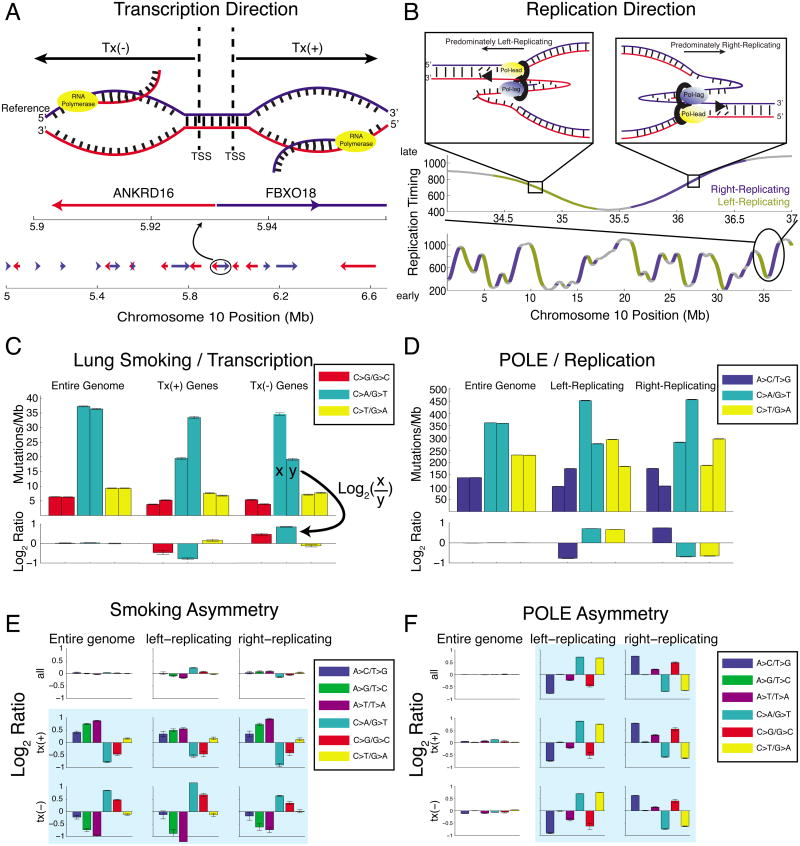
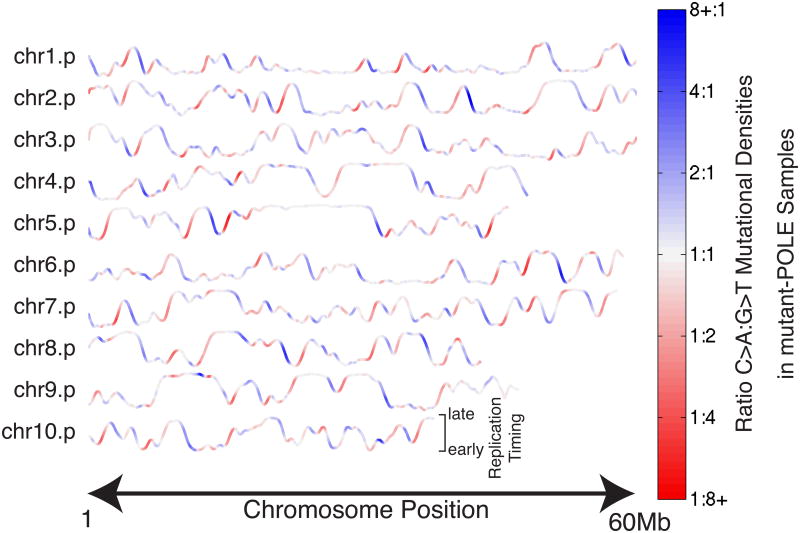
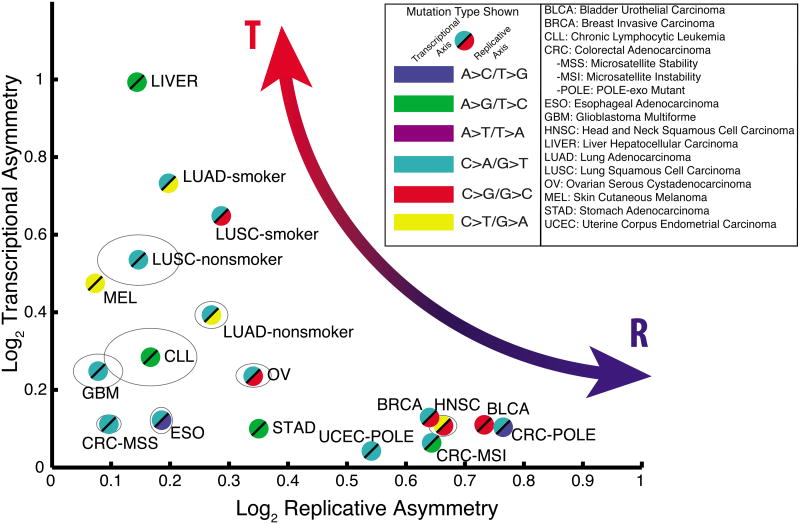
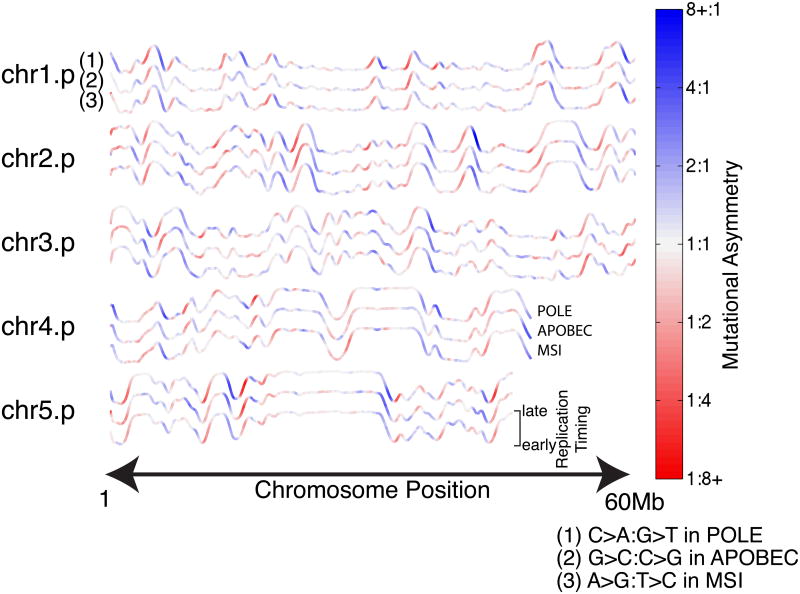
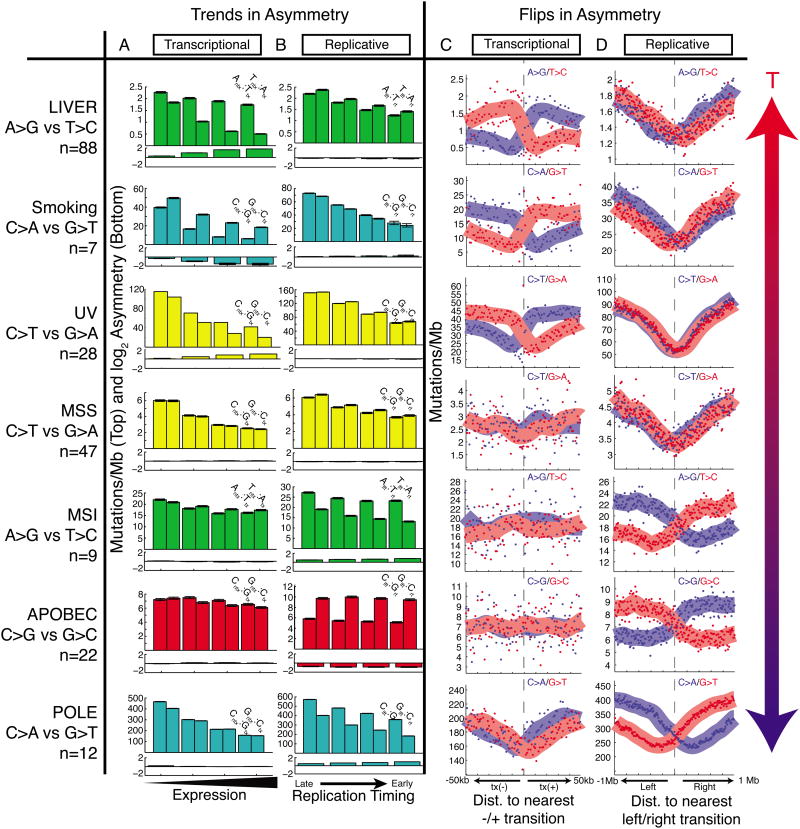
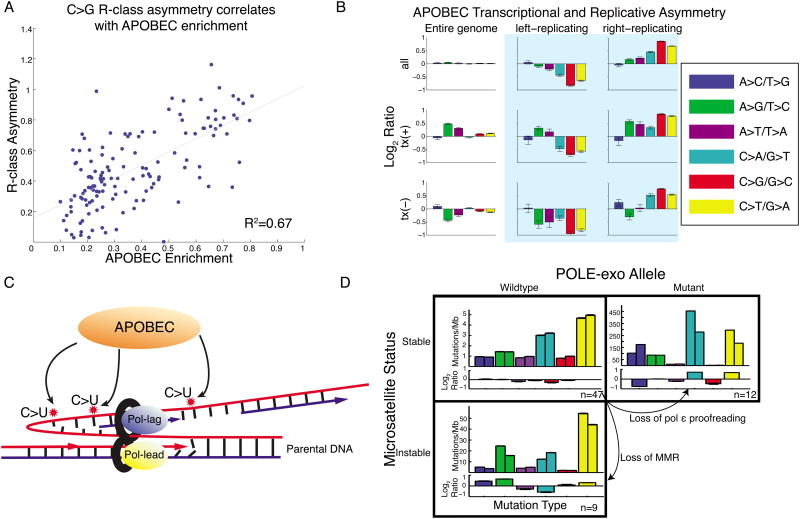
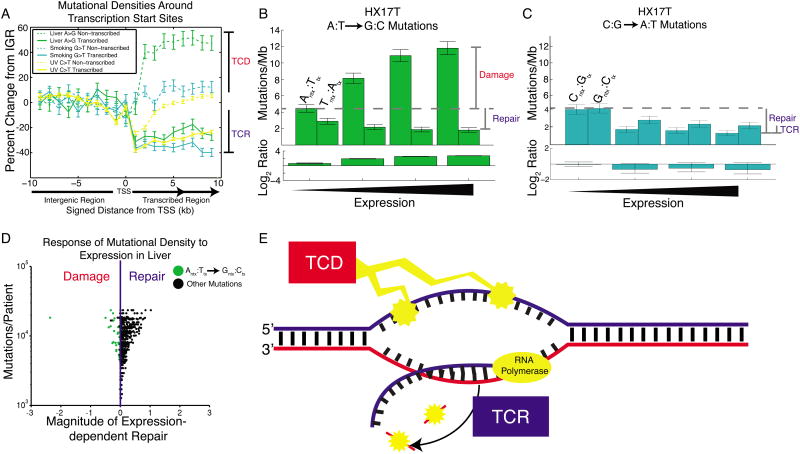
Mutational processes constantly shape the somatic genome, leading to immunity, aging, cancer, and other diseases. When cancer is the outcome, we are afforded a glimpse into these processes by the clonal expansion of the malignant cell. Here, we characterize a less explored layer of the mutational landscape of cancer: mutational asymmetries between the two DNA strands. Analyzing whole-genome sequences of 590 tumors from 14 different cancer types, we reveal widespread asymmetries across mutagenic processes, with transcriptional ("T-class") asymmetry dominating UV-, smoking-, and liver-cancer-associated mutations and replicative ("R-class") asymmetry dominating POLE-, APOBEC-, and MSI-associated mutations. We report a striking phenomenon of transcription-coupled damage (TCD) on the non-transcribed DNA strand and provide evidence that APOBEC mutagenesis occurs on the lagging-strand template during DNA replication. As more genomes are sequenced, studying and classifying their asymmetries will illuminate the underlying biological mechanisms of DNA damage and repair.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Carreras Puigvert J, Sanjiv K, Helleday T. Targeting DNA repair, DNA metabolism and replication stress as anti-cancer strategies. FEBS J 2015 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
