

A two-state activation mechanism controls the histone methyltransferase Suv39h1
- PMID: 26807716
- PMCID: PMC4876634
- DOI: 10.1038/nchembio.2008
A two-state activation mechanism controls the histone methyltransferase Suv39h1
Abstract
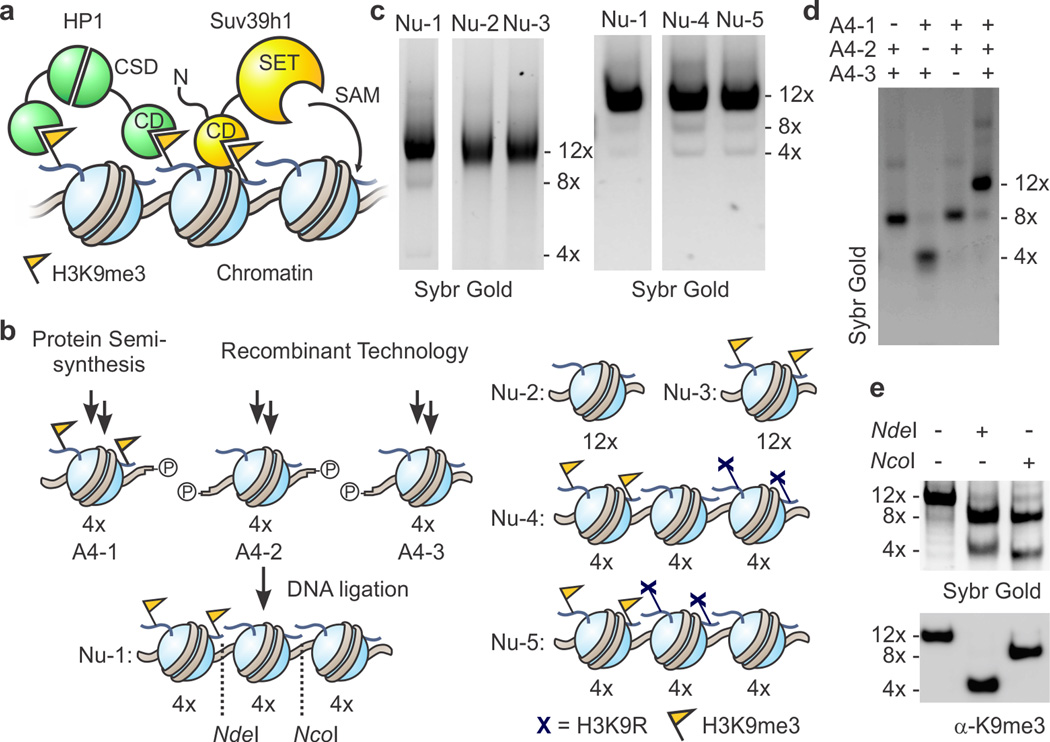
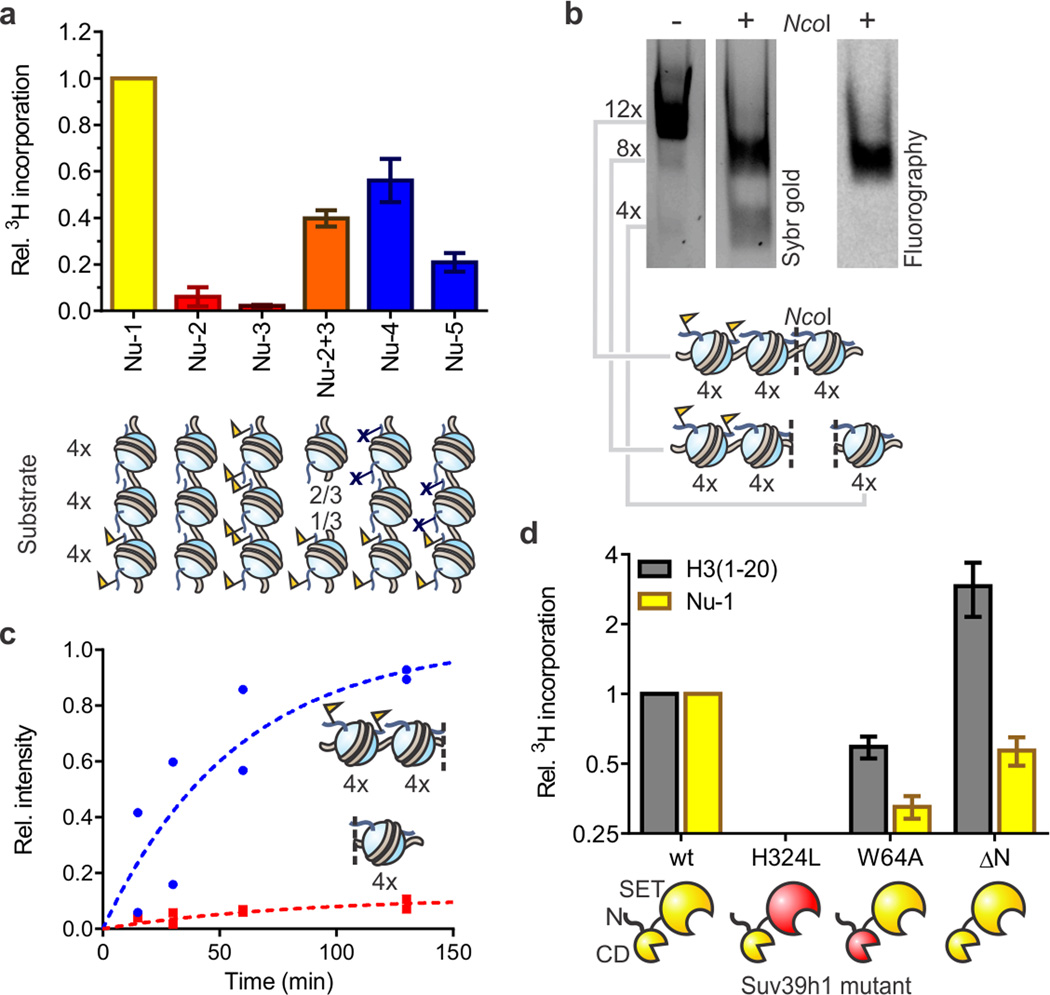
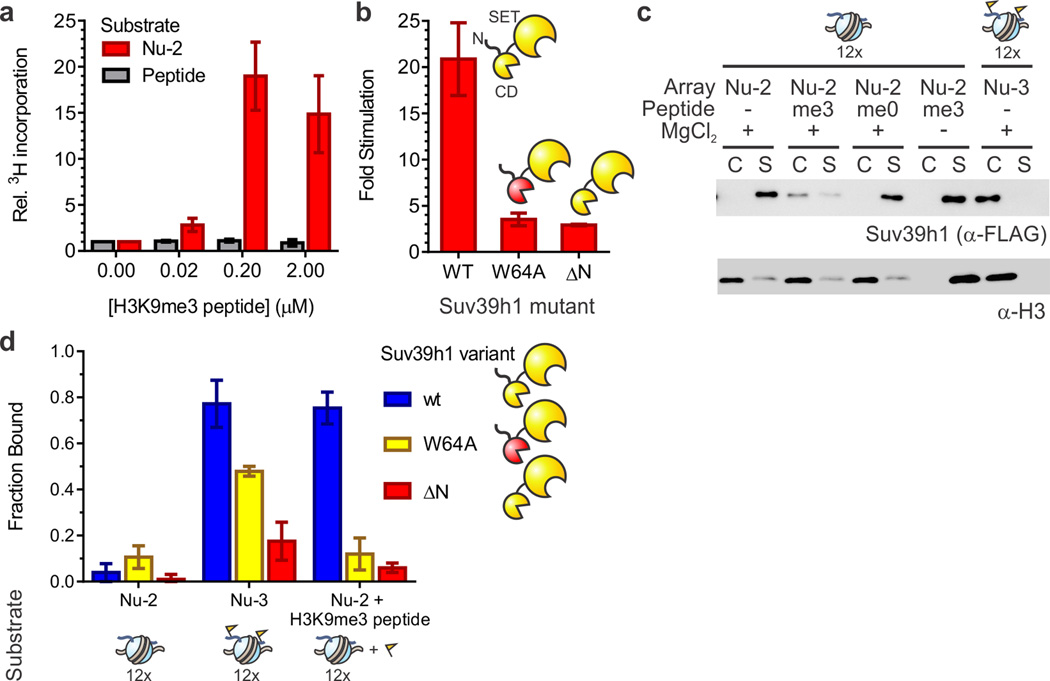
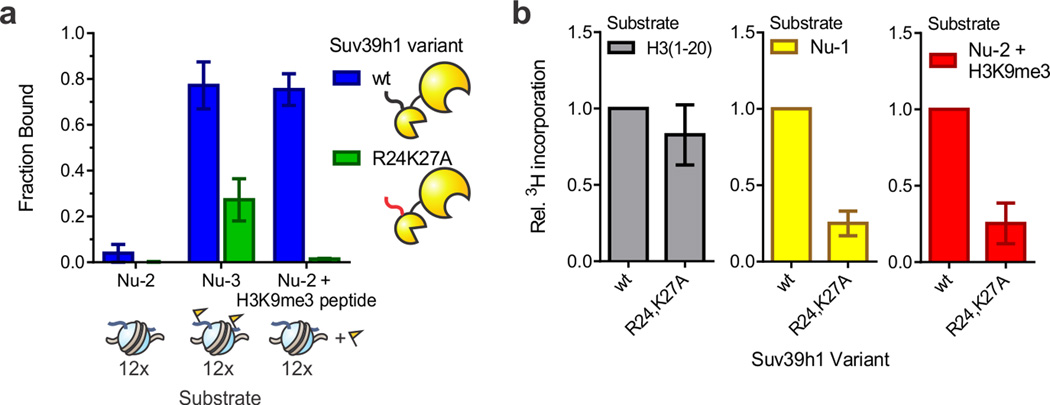
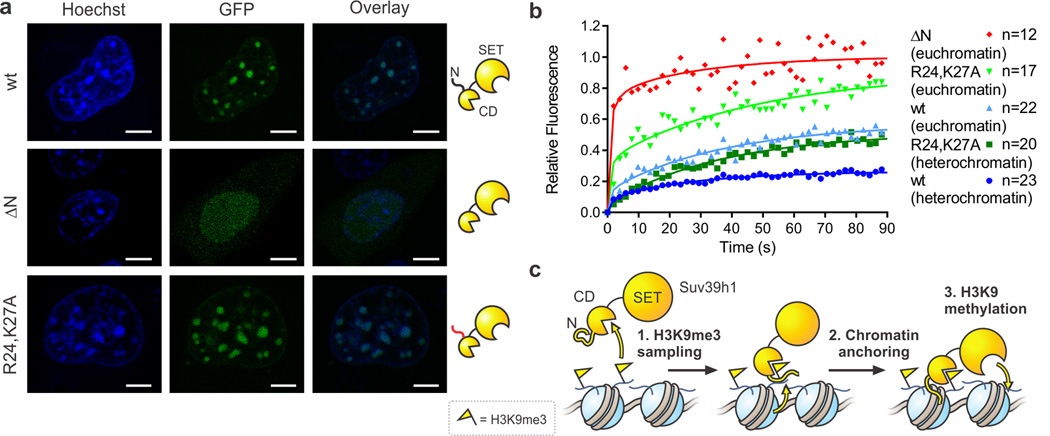
Specialized chromatin domains contribute to nuclear organization and regulation of gene expression. Gene-poor regions are di- and trimethylated at lysine 9 of histone H3 (H3K9me2 and H3K9me3) by the histone methyltransferase Suv39h1. This enzyme harnesses a positive feedback loop to spread H3K9me2 and H3K9me3 over extended heterochromatic regions. However, little is known about how feedback loops operate on complex biopolymers such as chromatin, in part because of the difficulty in obtaining suitable substrates. Here we describe the synthesis of multidomain 'designer chromatin' templates and their application to dissecting the regulation of human Suv39h1. We uncovered a two-step activation switch where H3K9me3 recognition and subsequent anchoring of the enzyme to chromatin allosterically promotes methylation activity and confirmed that this mechanism contributes to chromatin recognition in cells. We propose that this mechanism serves as a paradigm in chromatin biochemistry, as it enables highly dynamic sampling of chromatin state combined with targeted modification of desired genomic regions.
Figures
References
-
- Kornberg RD. Structure of chromatin. Annu Rev Biochem. 1977;46:931–954. - PubMed
-
- Rea S, et al. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature. 2000;406:593–599. - PubMed
-
- Nakayama J, Rice JC, Strahl BD, Allis CD, Grewal SI. Role of histone H3 lysine 9 methylation in epigenetic control of heterochromatin assembly. Science. 2001;292:110–113. - PubMed
-
- Grewal SI, Moazed D. Heterochromatin and epigenetic control of gene expression. Science. 2003;301:798–802. - PubMed
ONLINE METHODS REFERENCES
-
- Wan Q, Danishefsky SJ. Free-radical-based, specific desulfurization of cysteine: a powerful advance in the synthesis of polypeptides and glycopolypeptides. Angew Chem Int Ed. 2007;46:9248–9252. - PubMed
-
- Dyer PN, Edayathumangalam RS, White CL, Bao Y, Chakravarthy S, Muthurajan UM, Luger K. Reconstitution of nucleosome core particles from recombinant histones and DNA. Methods Enzymol. 2004;375:23–44. - PubMed
-
- Dorigo B, Schalch T, Bystricky K, Richmond TJ. Chromatin fiber folding: Requirement for the histone H4 N-terminal tail. J Mol Biol. 2003;327:85–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
