

Functional Characterization of Pseudomonas Contact Dependent Growth Inhibition (CDI) Systems
- PMID: 26808644
- PMCID: PMC4725963
- DOI: 10.1371/journal.pone.0147435
Functional Characterization of Pseudomonas Contact Dependent Growth Inhibition (CDI) Systems
Erratum in
-
Correction: Functional Characterization of Pseudomonas Contact Dependent Growth Inhibition (CDI) Systems.PLoS One. 2016 Feb 25;11(2):e0150538. doi: 10.1371/journal.pone.0150538. eCollection 2016. PLoS One. 2016. PMID: 26914642 Free PMC article. No abstract available.
Abstract
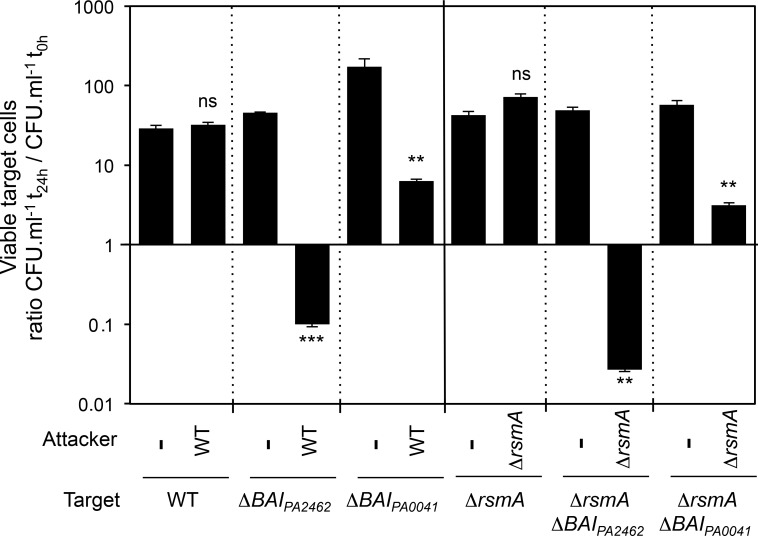
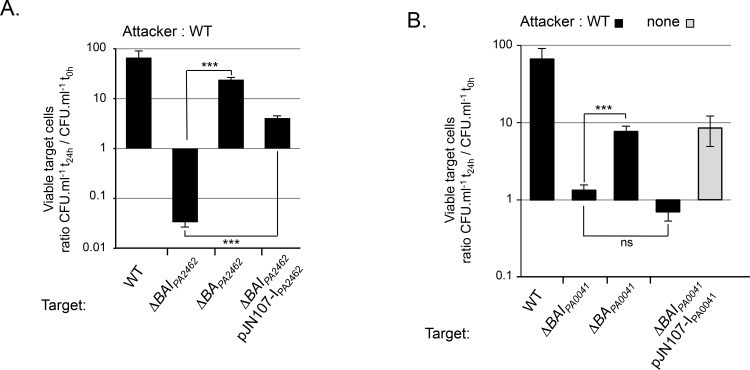
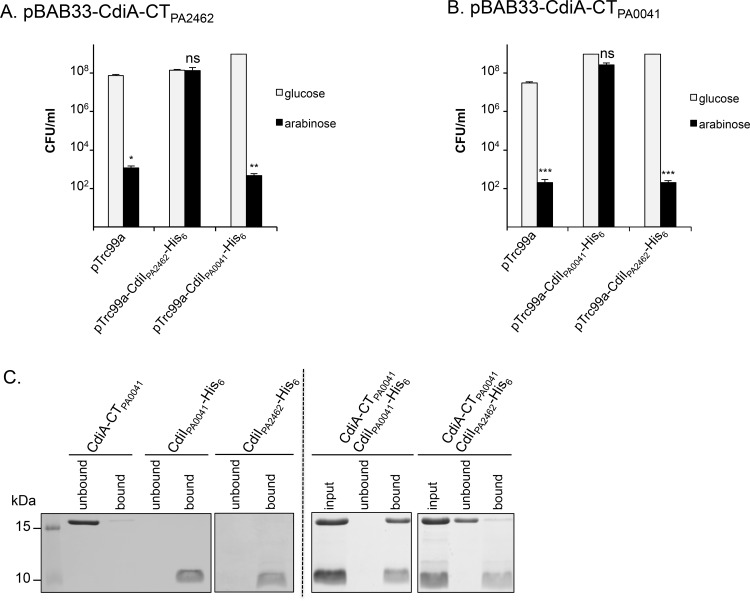
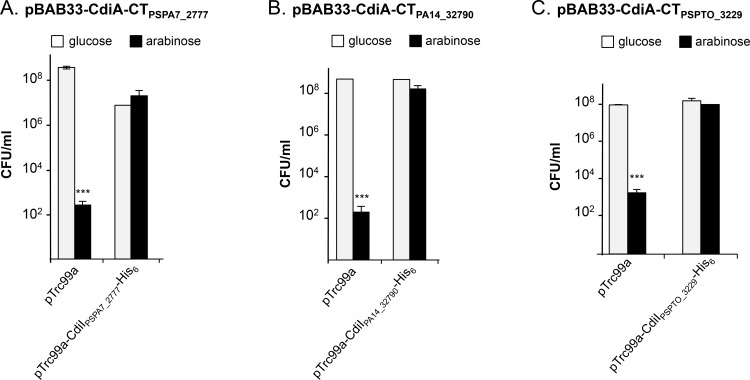
Contact-dependent inhibition (CDI) toxins, delivered into the cytoplasm of target bacterial cells, confer to host strain a significant competitive advantage. Upon cell contact, the toxic C-terminal region of surface-exposed CdiA protein (CdiA-CT) inhibits the growth of CDI- bacteria. CDI+ cells express a specific immunity protein, CdiI, which protects from autoinhibition by blocking the activity of cognate CdiA-CT. CdiA-CT are separated from the rest of the protein by conserved peptide motifs falling into two distinct classes, the "E. coli"- and "Burkholderia-type". CDI systems have been described in numerous species except in Pseudomonadaceae. In this study, we identified functional toxin/immunity genes linked to CDI systems in the Pseudomonas genus, which extend beyond the conventional CDI classes by the variability of the peptide motif that delimits the polymorphic CdiA-CT domain. Using P. aeruginosa PAO1 as a model, we identified the translational repressor RsmA as a negative regulator of CDI systems. Our data further suggest that under conditions of expression, P. aeruginosa CDI systems are implicated in adhesion and biofilm formation and provide an advantage in competition assays. All together our data imply that CDI systems could play an important role in niche adaptation of Pseudomonadaceae.
Conflict of interest statement
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
