

Makorin ortholog LEP-2 regulates LIN-28 stability to promote the juvenile-to-adult transition in Caenorhabditis elegans
- PMID: 26811380
- PMCID: PMC4813343
- DOI: 10.1242/dev.132738
Makorin ortholog LEP-2 regulates LIN-28 stability to promote the juvenile-to-adult transition in Caenorhabditis elegans
Abstract
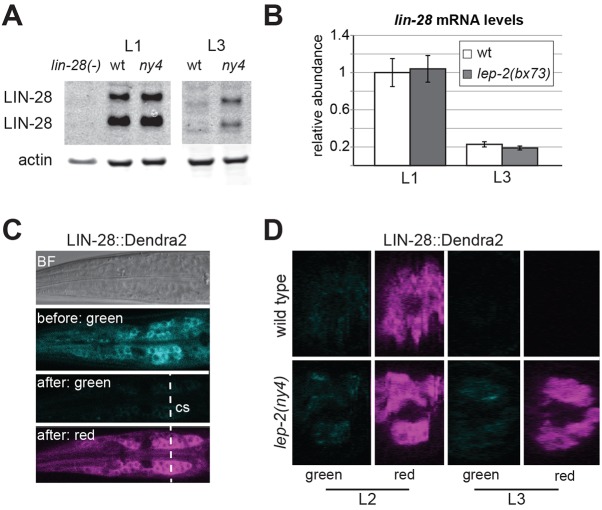
The heterochronic genes lin-28, let-7 and lin-41 regulate fundamental developmental transitions in animals, such as stemness versus differentiation and juvenile versus adult states. We identify a new heterochronic gene, lep-2, in Caenorhabditis elegans. Mutations in lep-2 cause a delay in the juvenile-to-adult transition, with adult males retaining pointed, juvenile tail tips, and displaying defective sexual behaviors. In both sexes, lep-2 mutants fail to cease molting or produce an adult cuticle. We find that LEP-2 post-translationally regulates LIN-28 by promoting LIN-28 protein degradation. lep-2 encodes the sole C. elegans ortholog of the Makorin (Mkrn) family of proteins. Like lin-28 and other heterochronic pathway members, vertebrate Mkrns are involved in developmental switches, including the timing of pubertal onset in humans. Based on shared roles, conservation and the interaction between lep-2 and lin-28 shown here, we propose that Mkrns, together with other heterochronic genes, constitute an evolutionarily ancient conserved module regulating switches in development.
Keywords: Developmental timing; Heterochronic pathway; LIN-28/let-7 axis; Mkrn.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
Similar articles
-
The Makorin lep-2 and the lncRNA lep-5 regulate lin-28 to schedule sexual maturation of the C. elegans nervous system.Elife. 2019 Jul 2;8:e43660. doi: 10.7554/eLife.43660. Elife. 2019. PMID: 31264582 Free PMC article.
-
The Long Non-Coding RNA lep-5 Promotes the Juvenile-to-Adult Transition by Destabilizing LIN-28.Dev Cell. 2019 May 20;49(4):542-555.e9. doi: 10.1016/j.devcel.2019.03.003. Epub 2019 Apr 4. Dev Cell. 2019. PMID: 30956008 Free PMC article.
-
Novel gain-of-function alleles demonstrate a role for the heterochronic gene lin-41 in C. elegans male tail tip morphogenesis.Dev Biol. 2006 Sep 1;297(1):74-86. doi: 10.1016/j.ydbio.2006.04.472. Epub 2006 May 6. Dev Biol. 2006. PMID: 16806150
-
Control of developmental timing by small temporal RNAs: a paradigm for RNA-mediated regulation of gene expression.Bioessays. 2002 Feb;24(2):119-29. doi: 10.1002/bies.10046. Bioessays. 2002. PMID: 11835276 Review.
-
LET-23-mediated signal transduction during Caenorhabditis elegans development.Mol Reprod Dev. 1995 Dec;42(4):523-8. doi: 10.1002/mrd.1080420422. Mol Reprod Dev. 1995. PMID: 8607985 Review.
Cited by
-
Tag team: Roles of miRNAs and Proteolytic Regulators in Ensuring Robust Gene Expression Dynamics.Trends Genet. 2018 Jan;34(1):21-29. doi: 10.1016/j.tig.2017.09.009. Epub 2017 Oct 13. Trends Genet. 2018. PMID: 29037438 Free PMC article. Review.
-
Sexual modulation of sex-shared neurons and circuits in Caenorhabditis elegans.J Neurosci Res. 2017 Jan 2;95(1-2):527-538. doi: 10.1002/jnr.23912. J Neurosci Res. 2017. PMID: 27870393 Free PMC article. Review.
-
The Makorin lep-2 and the lncRNA lep-5 regulate lin-28 to schedule sexual maturation of the C. elegans nervous system.Elife. 2019 Jul 2;8:e43660. doi: 10.7554/eLife.43660. Elife. 2019. PMID: 31264582 Free PMC article.
-
Ubiquitin ligases and a processive proteasome facilitate protein clearance during the oocyte-to-embryo transition in Caenorhabditis elegans.Genetics. 2022 May 5;221(1):iyac051. doi: 10.1093/genetics/iyac051. Genetics. 2022. PMID: 35377419 Free PMC article.
-
Neurogenesis in Caenorhabditis elegans.Genetics. 2024 Oct 7;228(2):iyae116. doi: 10.1093/genetics/iyae116. Genetics. 2024. PMID: 39167071 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
