

Transplant Antennae and Host Brain Interact to Shape Odor Perceptual Space in Male Moths
- PMID: 26816291
- PMCID: PMC4729490
- DOI: 10.1371/journal.pone.0147906
Transplant Antennae and Host Brain Interact to Shape Odor Perceptual Space in Male Moths
Abstract
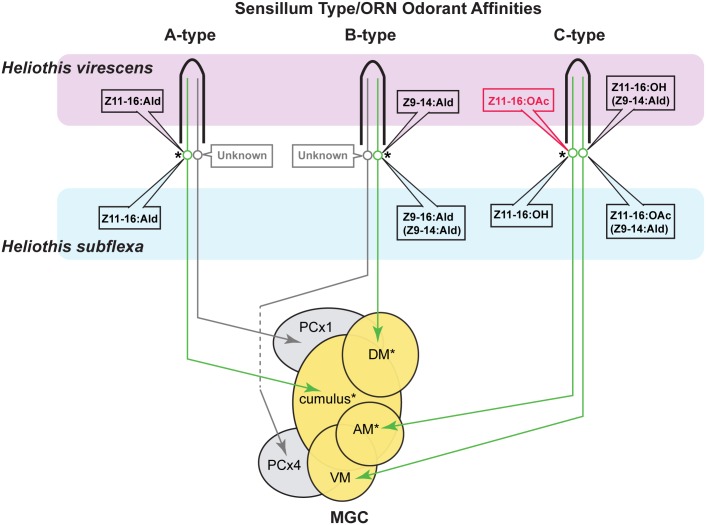
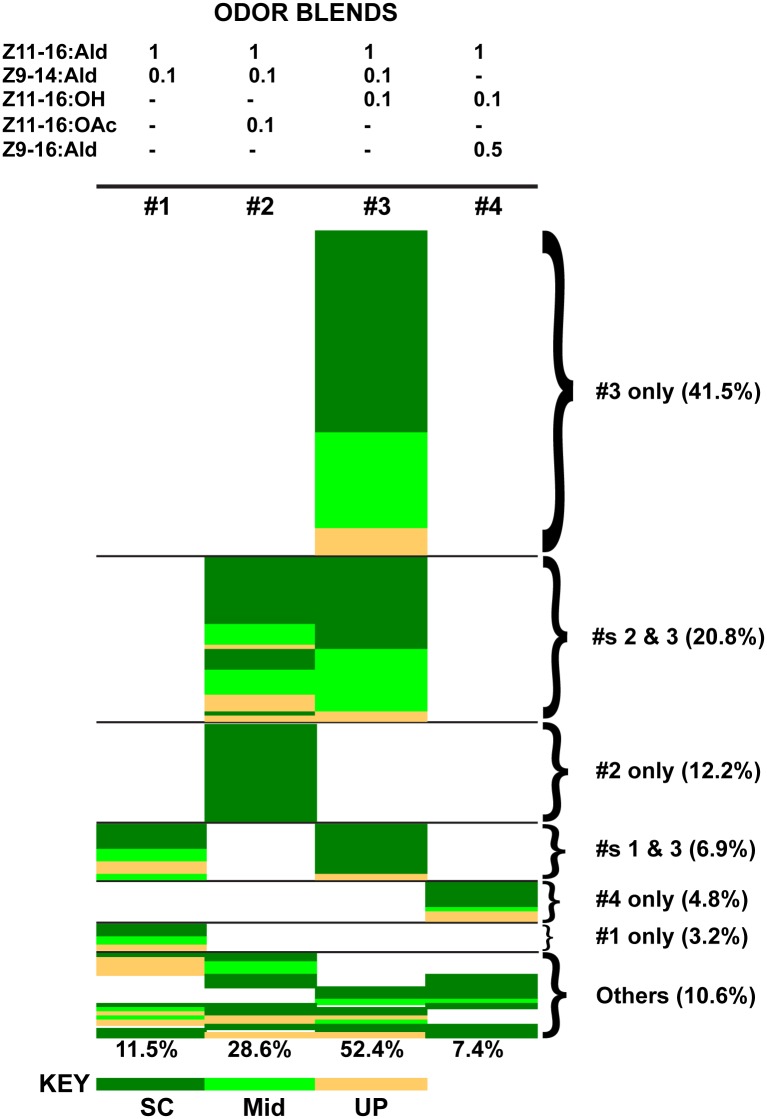
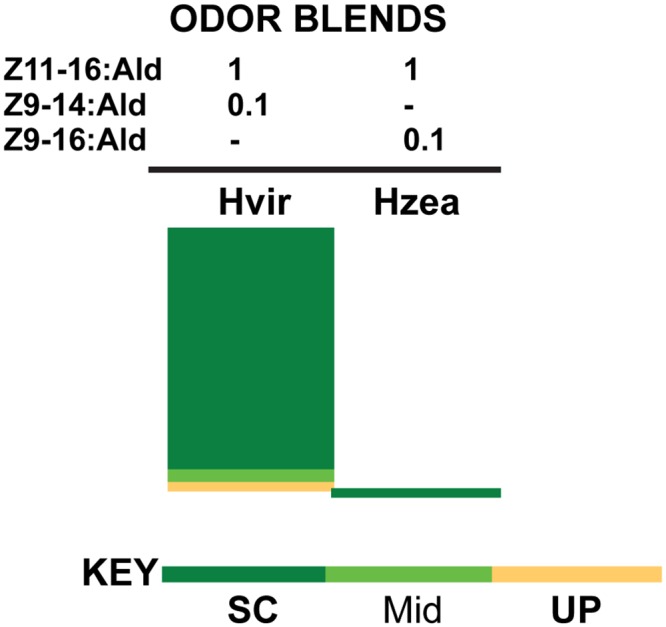
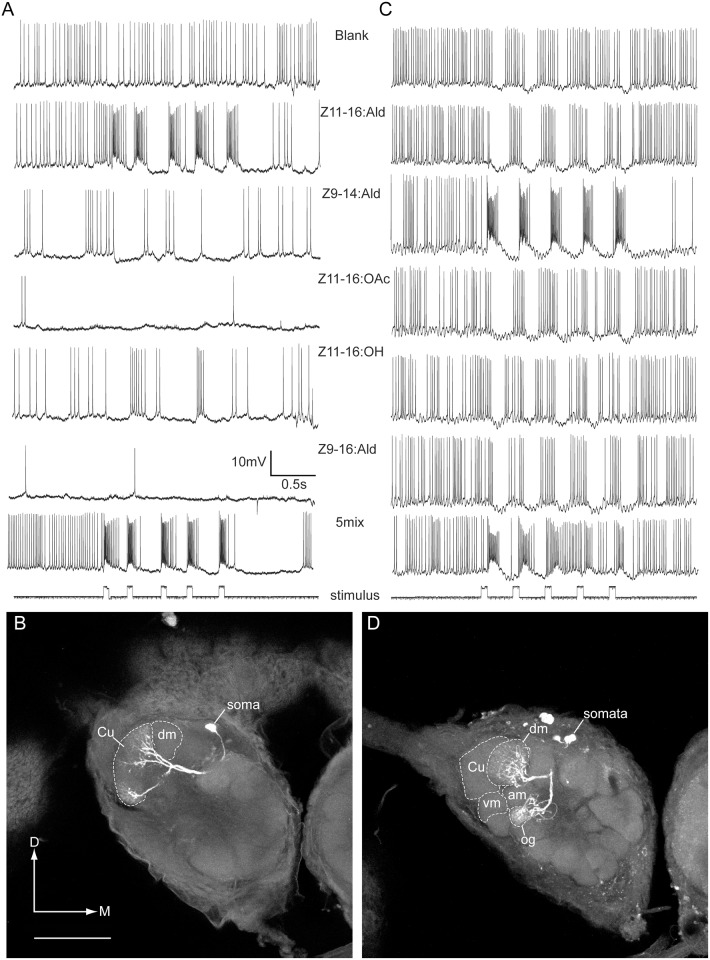
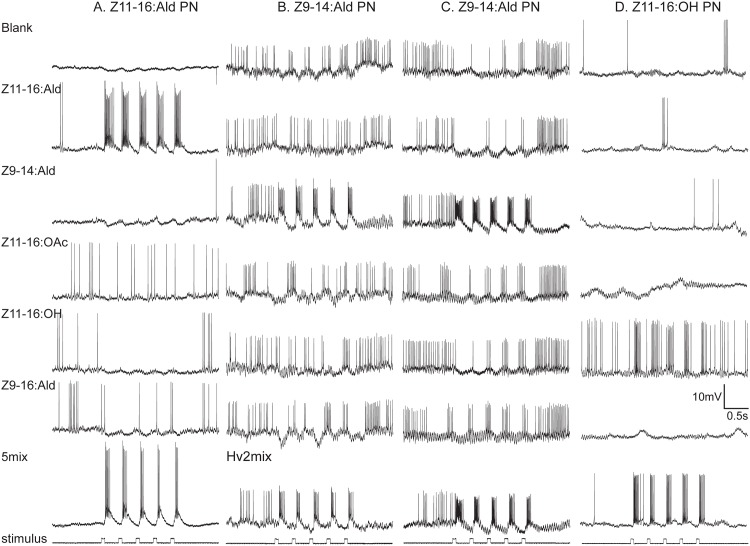
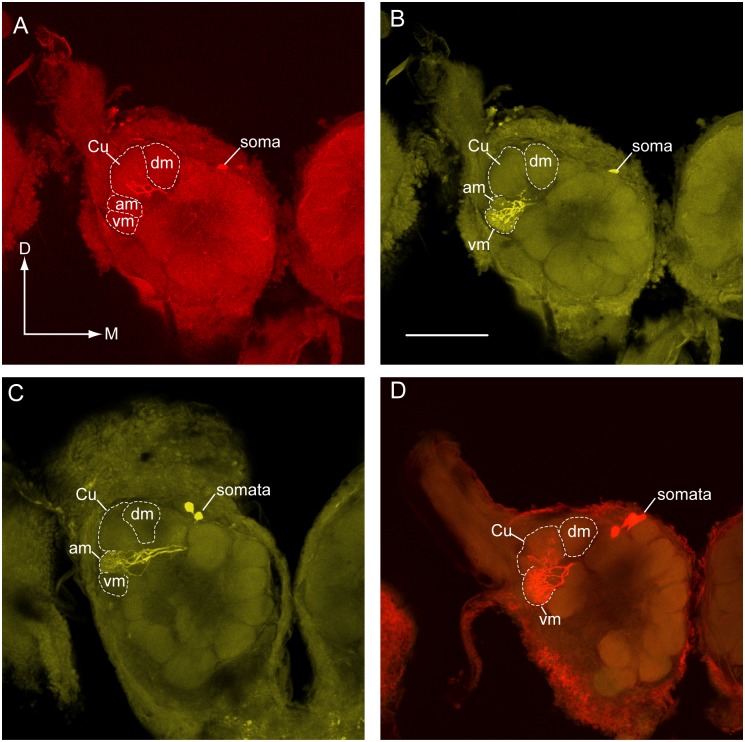
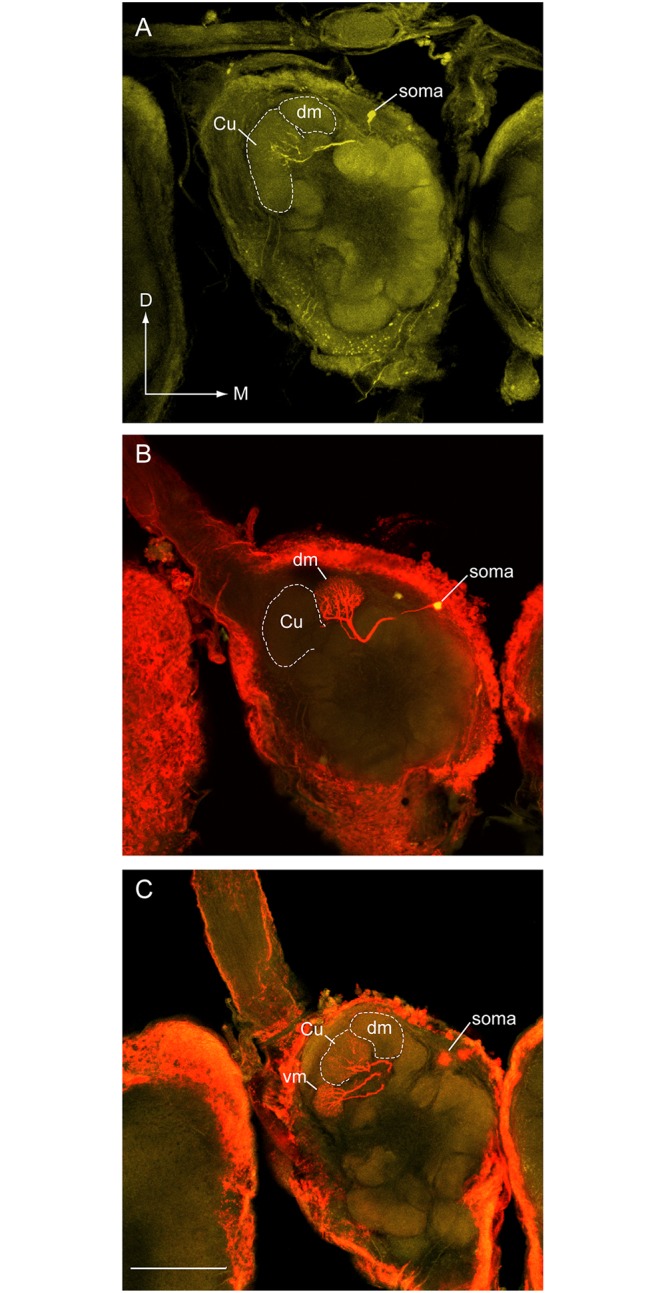
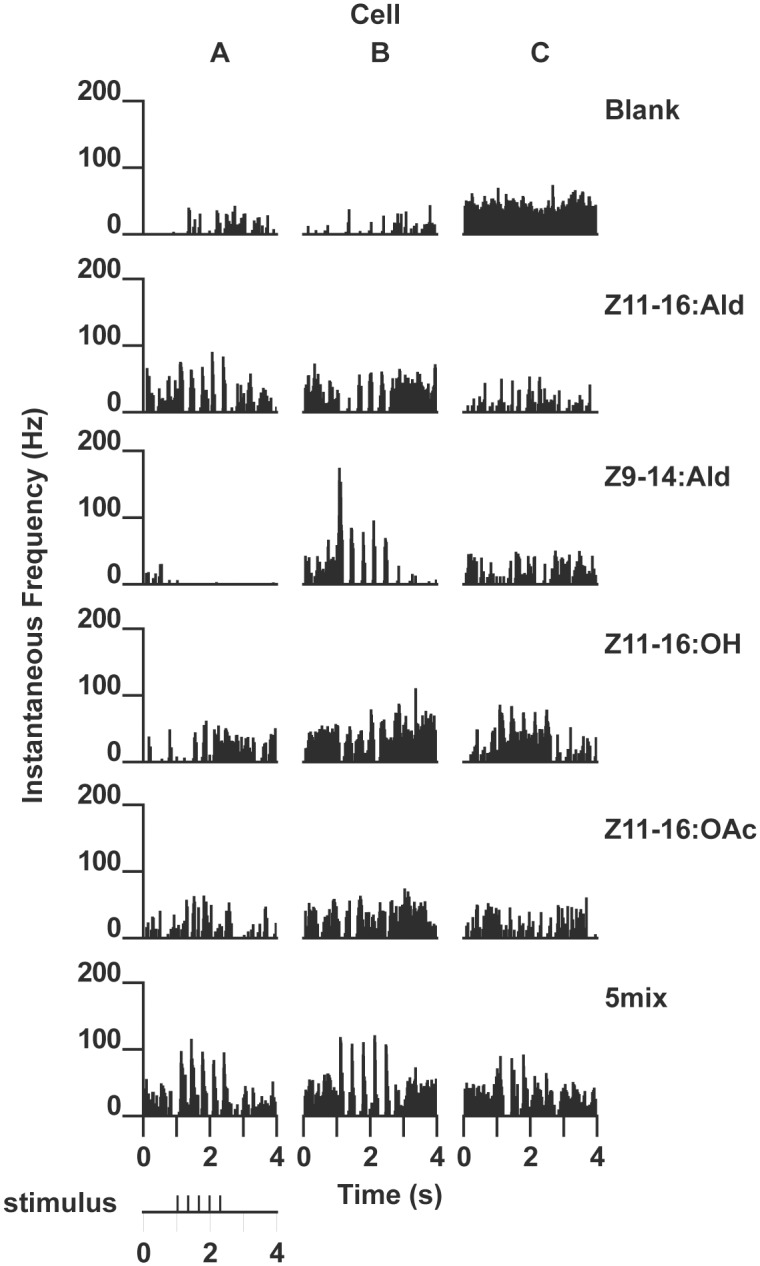
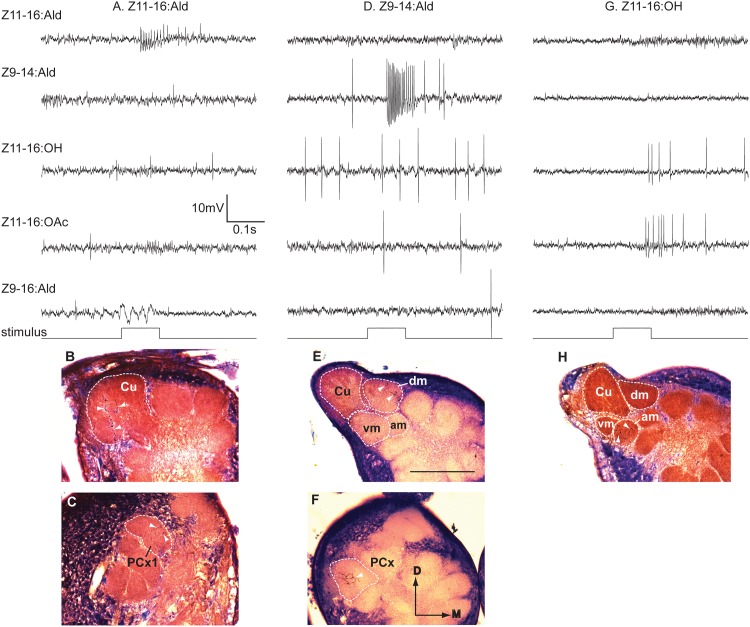
Behavioral responses to odors rely first upon their accurate detection by peripheral sensory organs followed by subsequent processing within the brain's olfactory system and higher centers. These processes allow the animal to form a unified impression of the odor environment and recognize combinations of odorants as single entities. To investigate how interactions between peripheral and central olfactory pathways shape odor perception, we transplanted antennal imaginal discs between larval males of two species of moth Heliothis virescens and Heliothis subflexa that utilize distinct pheromone blends. During metamorphic development olfactory receptor neurons originating from transplanted discs formed connections with host brain neurons within olfactory glomeruli of the adult antennal lobe. The normal antennal receptor repertoire exhibited by males of each species reflects the differences in the pheromone blends that these species employ. Behavioral assays of adult transplant males revealed high response levels to two odor blends that were dissimilar from those that attract normal males of either species. Neurophysiological analyses of peripheral receptor neurons and central olfactory neurons revealed that these behavioral responses were a result of: 1. the specificity of H. virescens donor olfactory receptor neurons for odorants unique to the donor pheromone blend and, 2. central odor recognition by the H. subflexa host brain, which typically requires peripheral receptor input across 3 distinct odor channels in order to elicit behavioral responses.
Conflict of interest statement
Figures
Similar articles
-
Inheritance of olfactory preferences III. Processing of pheromonal signals in the antennal lobe of Heliothis subflexa x Heliothis virescens hybrid male moths.Brain Behav Evol. 2006;68(2):90-108. doi: 10.1159/000093376. Epub 2006 May 16. Brain Behav Evol. 2006. PMID: 16707862
-
Inheritance of olfactory preferences II. Olfactory receptor neuron responses from Heliothis subflexa x Heliothis virescens hybrid male moths.Brain Behav Evol. 2006;68(2):75-89. doi: 10.1159/000093375. Epub 2006 May 16. Brain Behav Evol. 2006. PMID: 16707861
-
Moth pheromone-selective projection neurons with cell bodies in the antennal lobe lateral cluster exhibit diverse morphological and neurophysiological characteristics.J Comp Neurol. 2019 May 15;527(9):1443-1460. doi: 10.1002/cne.24611. Epub 2019 Feb 5. J Comp Neurol. 2019. PMID: 30723902
-
Morphology and physiology of the olfactory system of blood-feeding insects.J Physiol Paris. 2014 Apr-Jun;108(2-3):96-111. doi: 10.1016/j.jphysparis.2014.04.006. Epub 2014 May 15. J Physiol Paris. 2014. PMID: 24836537 Review.
-
Glomerular interactions in olfactory processing channels of the antennal lobes.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2013 Nov;199(11):929-46. doi: 10.1007/s00359-013-0842-6. Epub 2013 Jul 28. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2013. PMID: 23893248 Free PMC article. Review.
Cited by
-
Evolution of olfactory circuits in insects.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020 May;206(3):353-367. doi: 10.1007/s00359-020-01399-6. Epub 2020 Jan 27. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020. PMID: 31984441 Free PMC article. Review.
References
-
- Galizia CG, Szyszka P (2008) Olfactory coding in the insect brain: molecular receptive ranges, spatial and temporal coding. Ent Exp Appl 128: 81–92.
-
- Fitt GP (1989) The ecology of Heliothis species in relation to agroecosystems. Ann Rev Entomol 34: 17–52.
-
- Mitter C, Poole RW, Matthews M (1993) Biosystematics of the Heliothinae (Lepidoptera: Noctuidae). Ann Rev Entomol 38: 207–225.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
