

Aberrant Activation of p38 MAP Kinase-Dependent Innate Immune Responses Is Toxic to Caenorhabditis elegans
- PMID: 26818074
- PMCID: PMC4777117
- DOI: 10.1534/g3.115.025650
Aberrant Activation of p38 MAP Kinase-Dependent Innate Immune Responses Is Toxic to Caenorhabditis elegans
Abstract
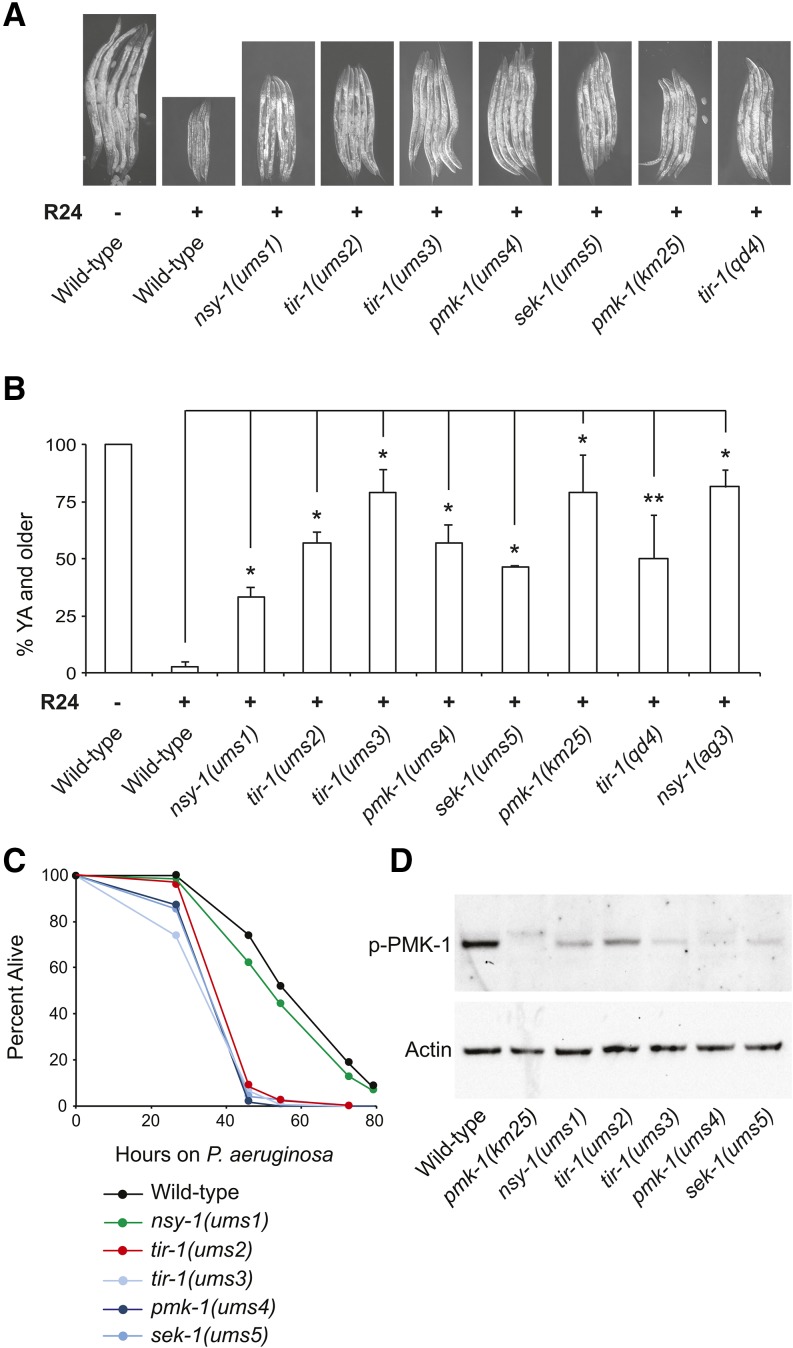
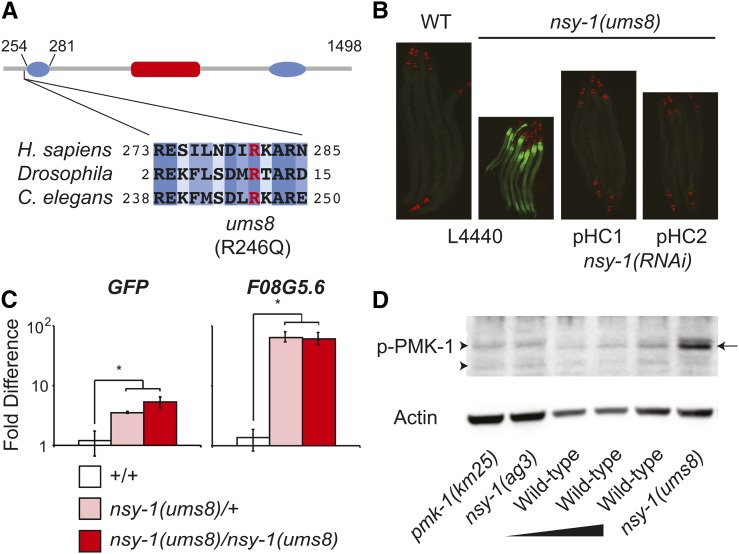
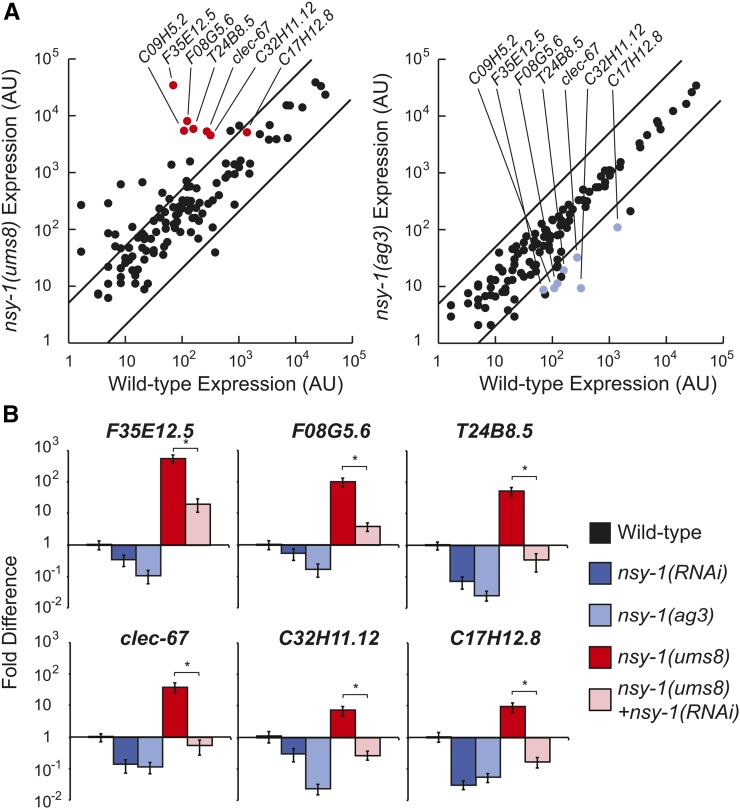
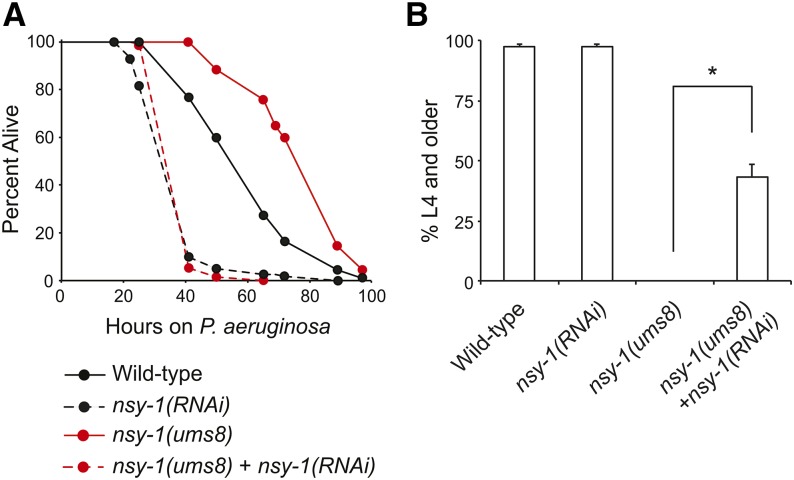
Inappropriate activation of innate immune responses in intestinal epithelial cells underlies the pathophysiology of inflammatory disorders of the intestine. Here we examine the physiological effects of immune hyperactivation in the intestine of the nematode Caenorhabditis elegans. We previously identified an immunostimulatory xenobiotic that protects C. elegans from bacterial infection by inducing immune effector expression via the conserved p38 MAP kinase pathway, but was toxic to nematodes developing in the absence of pathogen. To investigate a possible connection between the toxicity and immunostimulatory properties of this xenobiotic, we conducted a forward genetic screen for C. elegans mutants that are resistant to the deleterious effects of the compound, and identified five toxicity suppressors. These strains contained hypomorphic mutations in each of the known components of the p38 MAP kinase cassette (tir-1, nsy-1, sek-1, and pmk-1), demonstrating that hyperstimulation of the p38 MAPK pathway is toxic to animals. To explore mechanisms of immune pathway regulation in C. elegans, we conducted another genetic screen for dominant activators of the p38 MAPK pathway, and identified a single allele that had a gain-of-function (gf) mutation in nsy-1, the MAP kinase kinase kinase that acts upstream of p38 MAPK pmk-1. The nsy-1(gf) allele caused hyperinduction of p38 MAPK PMK-1-dependent immune effectors, had greater levels of phosphorylated p38 MAPK, and was more resistant to killing by the bacterial pathogen Pseudomonas aeruginosa compared to wild-type controls. In addition, the nsy-1(gf) mutation was toxic to developing animals. Together, these data suggest that the activity of the MAPKKK NSY-1 is tightly regulated as part of a physiological mechanism to control p38 MAPK-mediated innate immune hyperactivation, and ensure cellular homeostasis in C. elegans.
Keywords: C. elegans genetics; genetics of immunity; host-pathogen interactions; immune regulation; innate immunity.
Copyright © 2016 Cheesman et al.
Figures
Similar articles
-
A conserved p38 MAP kinase pathway in Caenorhabditis elegans innate immunity.Science. 2002 Jul 26;297(5581):623-6. doi: 10.1126/science.1073759. Science. 2002. PMID: 12142542
-
Early-life long-term exposure to ZnO nanoparticles suppresses innate immunity regulated by SKN-1/Nrf and the p38 MAPK signaling pathway in Caenorhabditis elegans.Environ Pollut. 2020 Jan;256:113382. doi: 10.1016/j.envpol.2019.113382. Epub 2019 Oct 16. Environ Pollut. 2020. PMID: 31662252
-
Caenorhabditis elegans mounts a p38 MAPK pathway-mediated defence to Cutibacterium acnes infection.Cell Microbiol. 2020 Oct;22(10):e13234. doi: 10.1111/cmi.13234. Epub 2020 Jul 15. Cell Microbiol. 2020. PMID: 32543022
-
Evolutionary perspectives on innate immunity from the study of Caenorhabditis elegans.Curr Opin Immunol. 2005 Feb;17(1):4-10. doi: 10.1016/j.coi.2004.11.007. Curr Opin Immunol. 2005. PMID: 15653303 Review.
-
Regulation of aging and innate immunity in C. elegans.Aging Cell. 2004 Aug;3(4):185-93. doi: 10.1111/j.1474-9728.2004.00108.x. Aging Cell. 2004. PMID: 15268752 Review.
Cited by
-
Non-canonical pattern recognition of a pathogen-derived metabolite by a nuclear hormone receptor identifies virulent bacteria in C. elegans.Immunity. 2023 Apr 11;56(4):768-782.e9. doi: 10.1016/j.immuni.2023.01.027. Epub 2023 Feb 17. Immunity. 2023. PMID: 36804958 Free PMC article.
-
Multiple pals gene modules control a balance between immunity and development in Caenorhabditis elegans.bioRxiv [Preprint]. 2023 Jan 18:2023.01.15.524171. doi: 10.1101/2023.01.15.524171. bioRxiv. 2023. Update in: PLoS Pathog. 2023 Jul 18;19(7):e1011120. doi: 10.1371/journal.ppat.1011120. PMID: 36711775 Free PMC article. Updated. Preprint.
-
Surveillance Immunity: An Emerging Paradigm of Innate Defense Activation in Caenorhabditis elegans.PLoS Pathog. 2016 Sep 15;12(9):e1005795. doi: 10.1371/journal.ppat.1005795. eCollection 2016 Sep. PLoS Pathog. 2016. PMID: 27631629 Free PMC article. Review. No abstract available.
-
Caenorhabditis elegans in high-throughput screens for anti-infective compounds.Curr Opin Immunol. 2018 Oct;54:59-65. doi: 10.1016/j.coi.2018.06.003. Epub 2018 Jun 20. Curr Opin Immunol. 2018. PMID: 29935375 Free PMC article. Review.
-
A transcriptome study on Macrobrachium nipponense hepatopancreas experimentally challenged with white spot syndrome virus (WSSV).PLoS One. 2018 Jul 6;13(7):e0200222. doi: 10.1371/journal.pone.0200222. eCollection 2018. PLoS One. 2018. PMID: 29979781 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous