

Progesterone modulates endothelial progenitor cell (EPC) viability through the CXCL12/CXCR4/PI3K/Akt signalling pathway
- PMID: 26818151
- PMCID: PMC6495665
- DOI: 10.1111/cpr.12231
Progesterone modulates endothelial progenitor cell (EPC) viability through the CXCL12/CXCR4/PI3K/Akt signalling pathway
Abstract
Objectives: Progesterone treatment can effectively increase levels of circulating endothelial progenitor cells (EPCs) and improve neurological functional outcome in a traumatic brain injury (TBI) rat model. However, the mechanisms of progesterone's effects on EPC viability remain elusive. The CXCL12/CXCR4 (CXC chemokine ligand 12/CXC chemokine receptor 4) signalling pathway regulates cell proliferation; we hypothesize that it mediates progesterone-induced EPC viability.
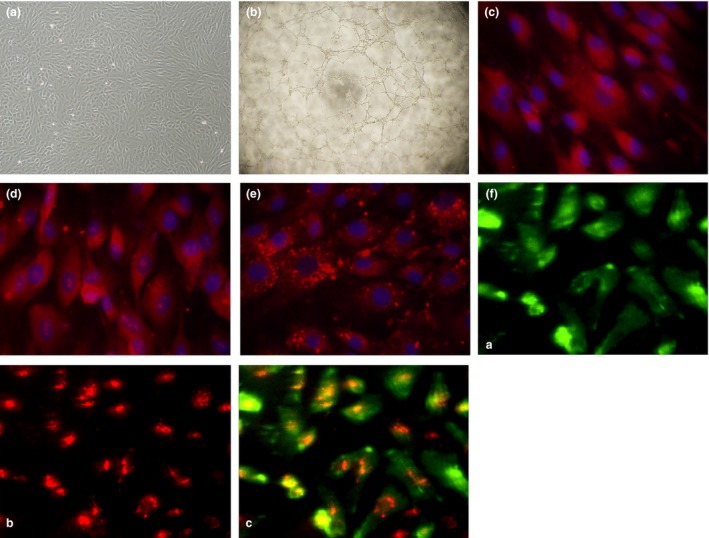
Materials and methods: EPCs were isolated from bone marrow-derived mononuclear cells (BM-MNCs) and treated with progesterone (5, 10 and 100 nm). MTS assay was used to investigate EPC viability. Protein expression was examined by Western blotting, ELISA assay and flow cytometry. Cell membrane and cytoplasm proteins were extracted with membrane and cytoplasm protein extraction kits. CXCR4 antagonist (AMD3100) and phosphatidylinositol 3-kinases (PI3K) antagonist (LY294002) were used to characterize underlying mechanisms.
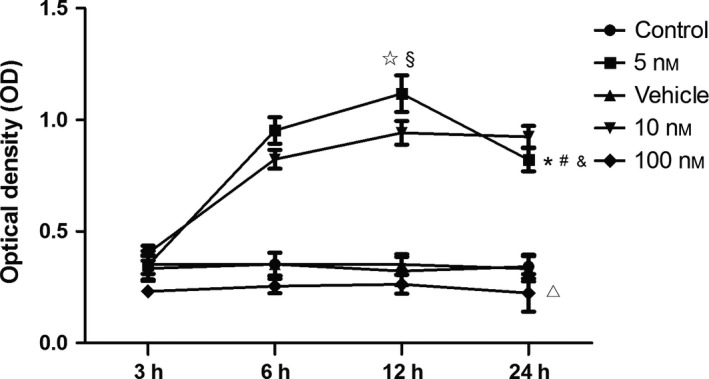
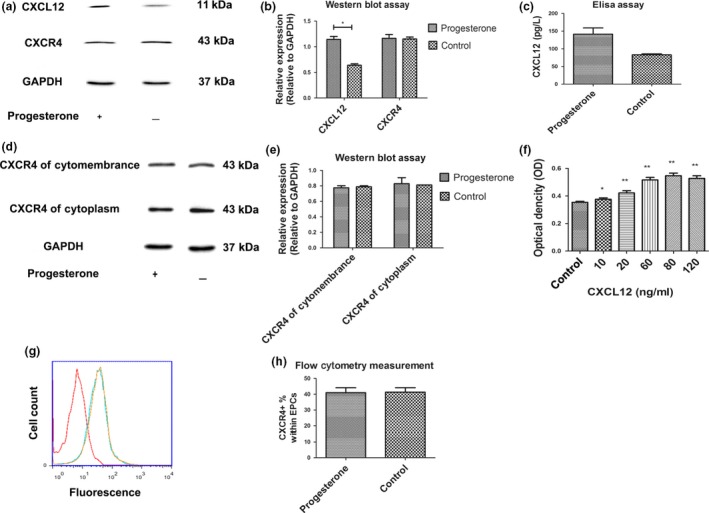
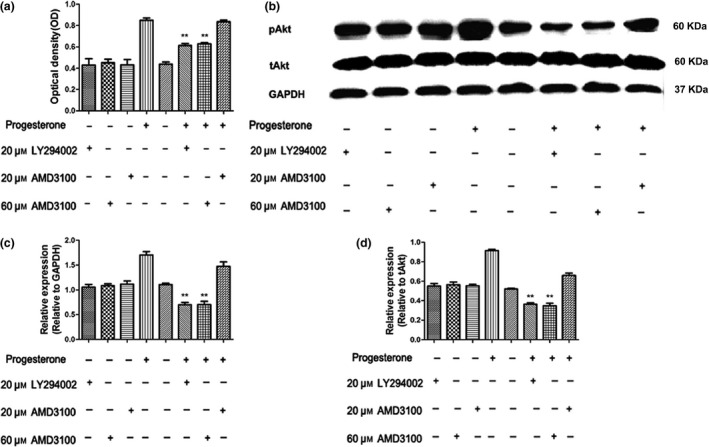
Results: Progesterone-induced EPC viability was time- and dose-dependent. Administration of progesterone facilitated EPC viability and increased expression of CXCL12 and phosphorylated Akt (also known as protein kinase B, pAkt) activity (P < 0.05). Progesterone did not regulate CXCR4 protein expression in cultured EPC membranes or cytoplasm. However, progesterone-induced EPC viability was significantly attenuated by AMD3100 or LY294002. Inhibition of the signalling pathway with AMD3100 and LY294002 subsequently reduced progesterone-induced CXCL12/CXCR4/PI3K/pAkt signalling activity.
Conclusions: The CXCL12/CXCR4/PI3K/pAkt signalling pathway increased progesterone-induced EPC viability.
© 2016 John Wiley & Sons Ltd.
Figures
References
-
- Hu Z, Li Y, Fang M, Wai MS, Yew DT (2009) Exogenous progesterone: a potential therapeutic candidate in CNS injury and neurodegeneration. Curr. Med. Chem. 16, 1418–1425. - PubMed
-
- Peluso JJ, Liu X, Saunders MM, Claffey KP, Phoenix K (2008) Regulation of ovarian cancer cell viability and sensitivity to cisplatin by progesterone receptor membrane component‐1. J. Clin. Endocrinol. Metab. 93, 1592–1599. - PubMed
-
- Peluso JJ, Yuan A, Liu X, Lodde V (2013) Plasminogen activator inhibitor 1 RNA‐binding protein interacts with progesterone receptor membrane component 1 to regulate progesterone's ability to maintain the viability of spontaneously immortalized granulosa cells and rat granulosa cells. Biol. Reprod. 88, 20. - PMC - PubMed
-
- Xue S, Zhang HT, Zhang P, Luo J, Chen ZZ, Jang XD et al (2010) Functional endothelial progenitor cells derived from adipose tissue show beneficial effect on cell therapy of traumatic brain injury. Neurosci. Lett. 473, 186–191. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
