

Cellular microenvironment controls the nuclear architecture of breast epithelia through β1-integrin
- PMID: 26818565
- PMCID: PMC4943696
- DOI: 10.1080/15384101.2015.1121354
Cellular microenvironment controls the nuclear architecture of breast epithelia through β1-integrin
Abstract
Defects in nuclear architecture occur in a variety of diseases, however the fundamental mechanisms that control the internal structure of nuclei are poorly defined. Here we reveal that the cellular microenvironment has a profound influence on the global internal organization of nuclei in breast epithelia. A 3D microenvironment induces a prolonged but reversible form of cell cycle arrest that features many of the classical markers of cell senescence. This unique form of arrest is dependent on signaling from the external microenvironment through β1-integrins. It is concomitant with alterations in nuclear architecture that characterize the withdrawal from cell proliferation. Unexpectedly, following prolonged cell cycle arrest in 3D, the senescence-like state and associated reprogramming of nuclear architecture are freely reversible on altering the dimensionality of the cellular microenvironment. Breast epithelia can therefore maintain a proliferative plasticity that correlates with nuclear remodelling. However, the changes in nuclear architecture are cell lineage-specific and do not occur in fibroblasts, and moreover they are overcome in breast cancer cells.
Keywords: breast cancer; breast mammary gland; cell cycle; cell senescence; extracellular matrix; integrin; nuclear structure.
Figures
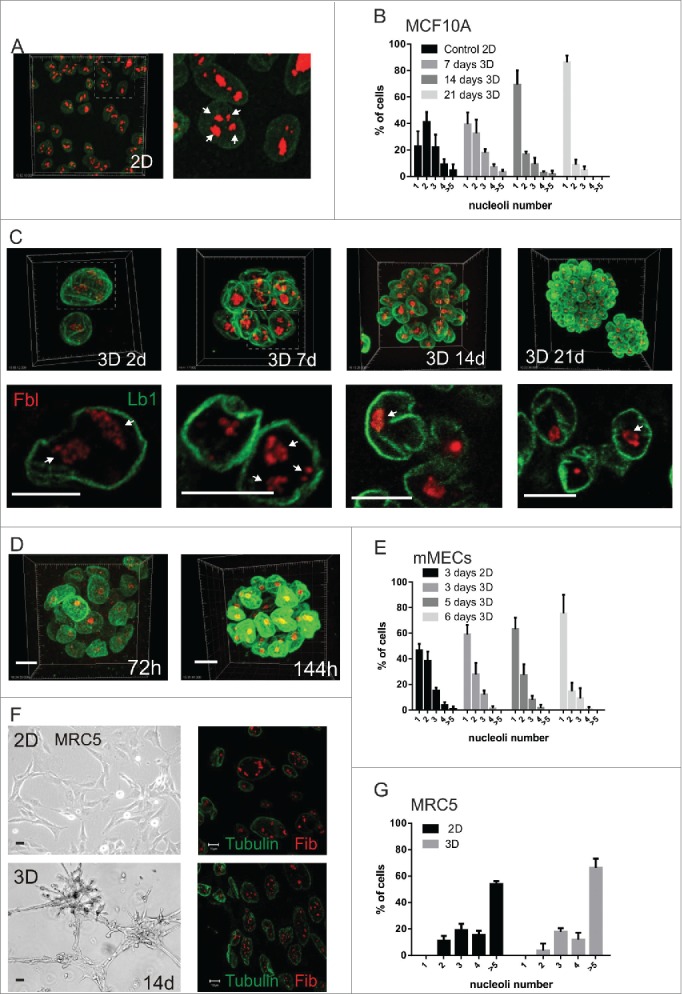
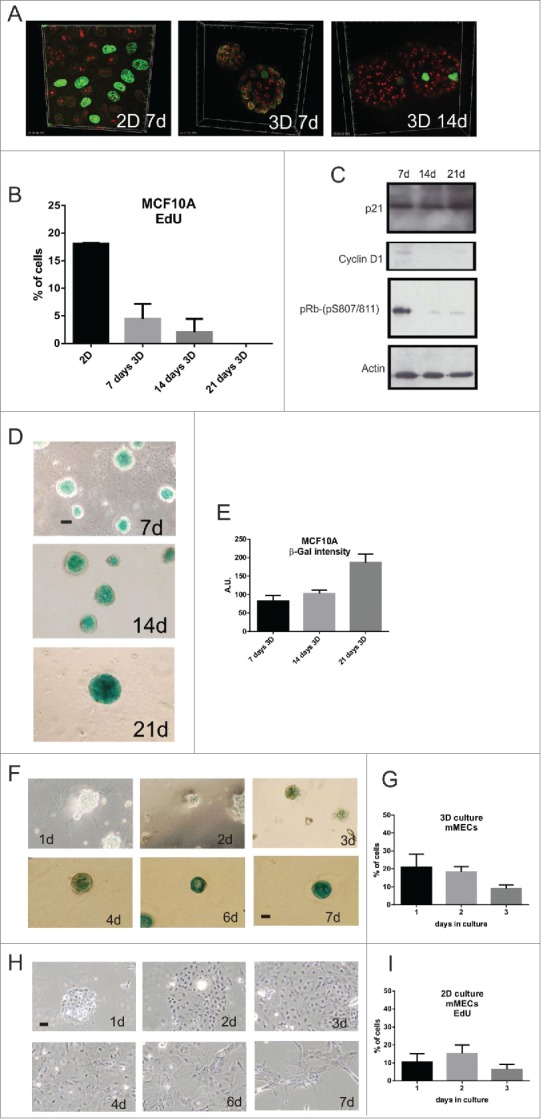
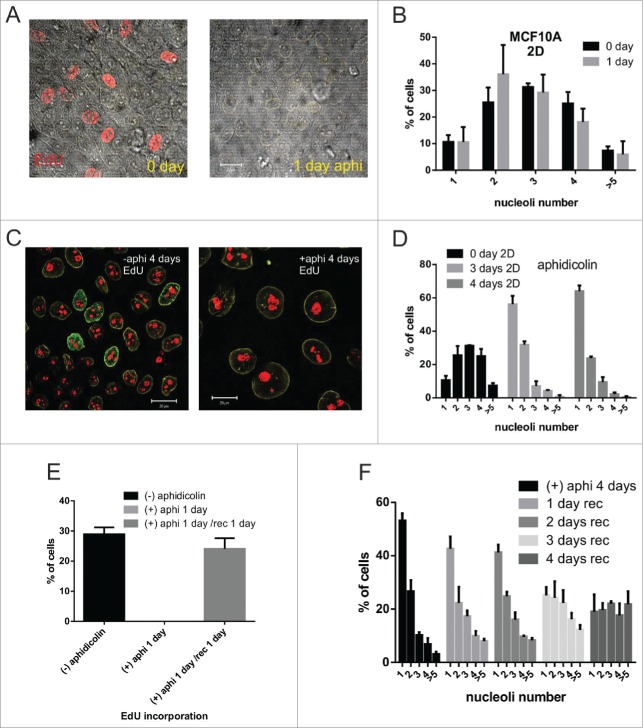
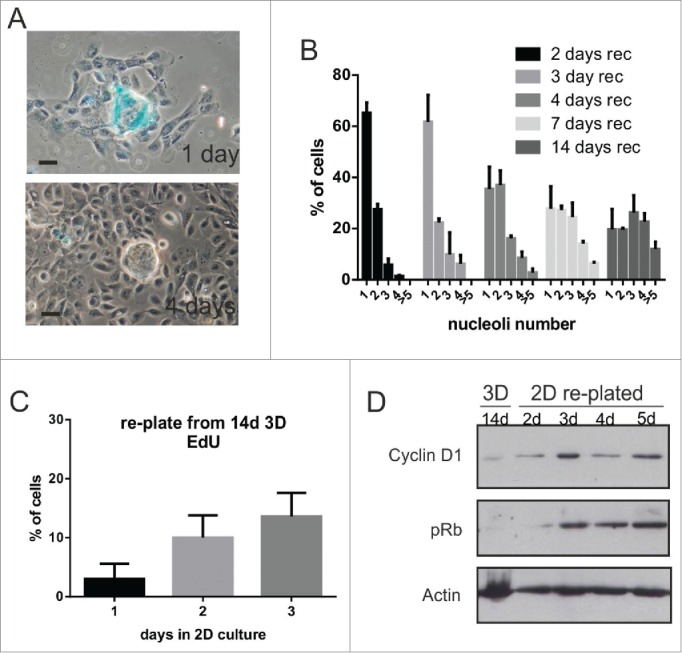
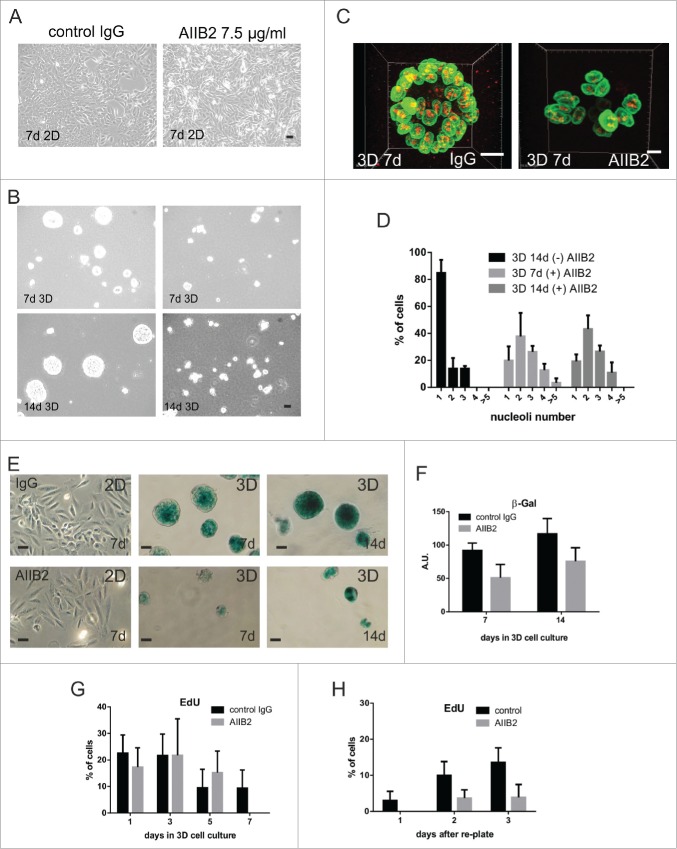
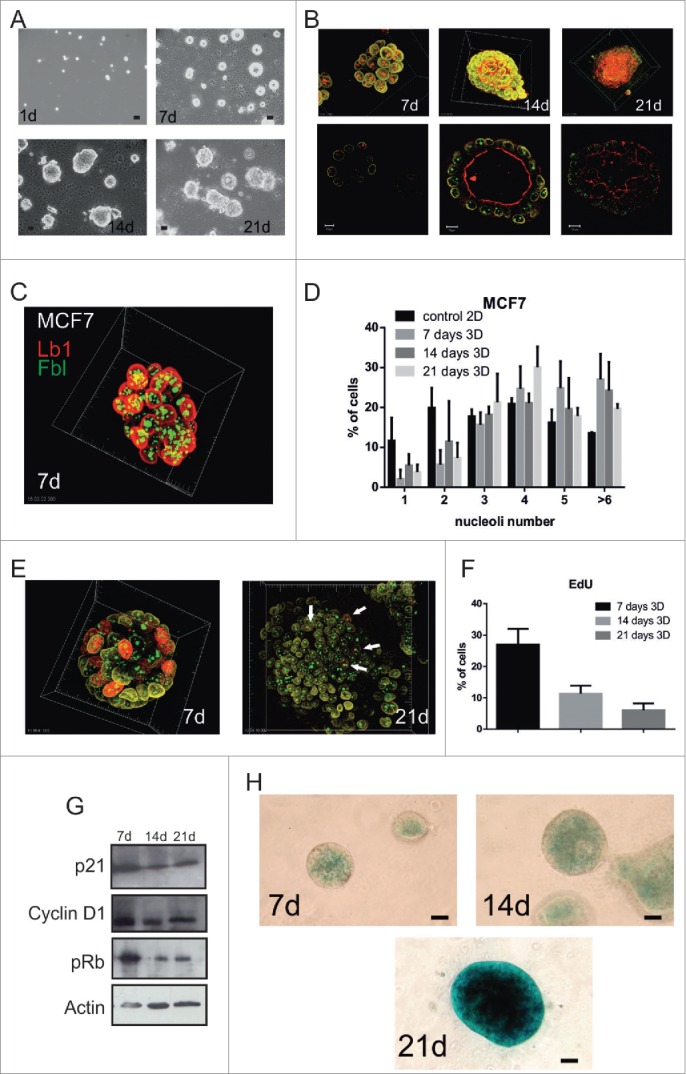
Comment in
-
A structured environment helps to regulate nuclear architecture in breast epithelial cells.Cell Cycle. 2016 May 2;15(9):1178-9. doi: 10.1080/15384101.2016.1147114. Epub 2016 Apr 14. Cell Cycle. 2016. PMID: 27078818 Free PMC article. No abstract available.
References
-
- Jackson DA. The principles of nuclear structure. Chromosome Res 2003; 11:387-401; PMID:12971716; http://dx.doi.org/ 10.1023/A:1024954123092 - DOI - PubMed
-
- Lamond AI, Spector DL. Nuclear speckles: a model for nuclear organelles. Nat Rev Mol Cell Biol 2003; 4:605-12; PMID:12923522; http://dx.doi.org/ 10.1038/nrm1172 - DOI - PubMed
-
- Handwerger KE, Gall JG. Subnuclear organelles: new insights into form and function. Trends Cell Biol 2006; 16:19-26; PMID:16325406; http://dx.doi.org/ 10.1016/j.tcb.2005.11.005 - DOI - PubMed
-
- Boisvert FM, van Koningsbruggen S, Navascues J, Lamond AI. The multifunctional nucleolus. Nat Rev Mol Cell Biol 2007; 8:574-85; PMID:17519961; http://dx.doi.org/ 10.1038/nrm2184 - DOI - PubMed
-
- Lanctot C, Cheutin T, Cremer M, Cavalli G, Cremer T. Dynamic genome architecture in the nuclear space: regulation of gene expression in three dimensions. Nat Rev Genet 2007; 8:104-15; PMID:17230197; http://dx.doi.org/ 10.1038/nrg2041 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources