

It takes a seasoned bird to be a good listener: communication between the sexes
- PMID: 26820470
- PMCID: PMC4921280
- DOI: 10.1016/j.conb.2016.01.005
It takes a seasoned bird to be a good listener: communication between the sexes
Abstract
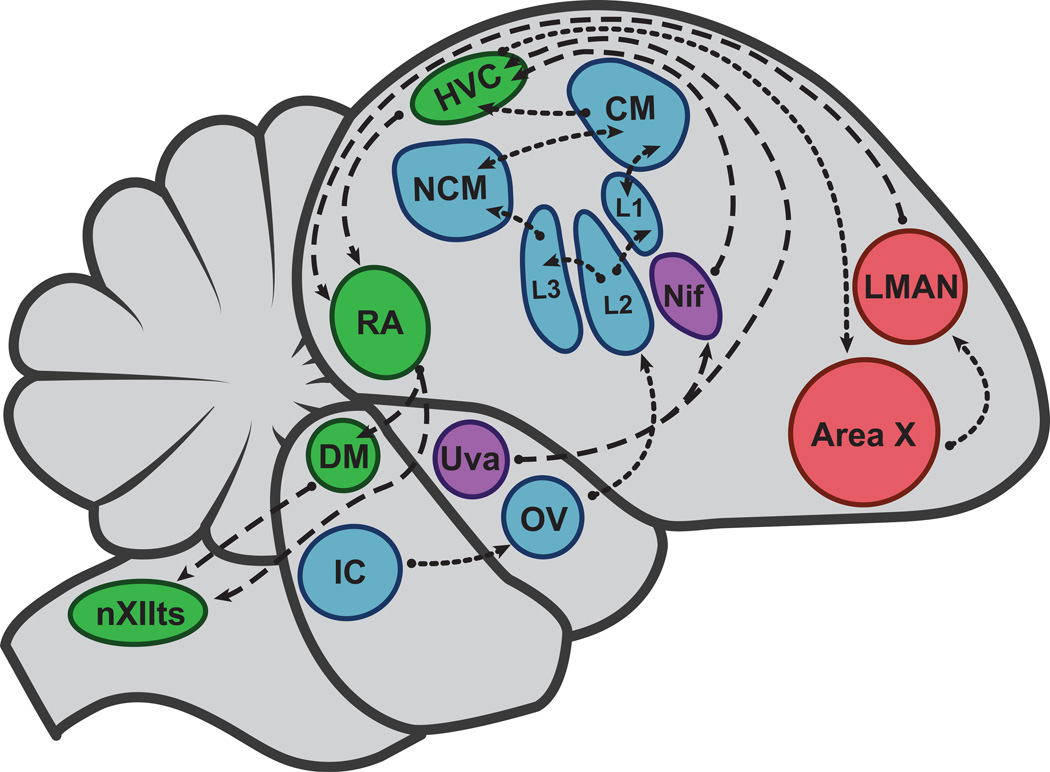
Birds commonly use sound for communication between the sexes. In many songbird species, only males sing and there are pronounced sex differences in the neural song control circuits. By contrast, the auditory circuitry is largely similar in males and females. Both sexes learn to recognize vocalizations heard as juveniles and this shapes auditory response selectivity. Mating vocalizations are restricted to the breeding season, when sex steroid levels are elevated. Auditory cells, from the ear to the cortex, are hormone sensitive. Estrogens are synthesized in the brain and can modulate the activity of auditory neurons. In species that breed seasonally, elevated levels of estradiol in females transiently enhance their auditory responses to conspecific vocalizations, resulting in sex differences in audition.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Beecher MD, Brenowitz EA. Functional aspects of song learning in the songbirds. Trends Ecol. Evol. 2005;20:143–149. - PubMed
-
- Brenowitz EA, Beecher MD. Song learning in birds: diversity and plasticity, opportunities and challenges. Trends Neurosci. 2005;28:127–132. - PubMed
-
-
Fortune ES, Rodríguez C, Li D, Ball GF, Coleman MJ. Neural Mechanisms for the Coordination of Duet Singing in Wrens. Science. 2011;334:666–670. Auditory activity in HVC was recorded from wild-caught male and female Bay Wrens, a neotropical species in which both sexes sing duets. Neurons in both sexes responded best to the combined male and female duet than to either sex's notes alone.
-
-
- Catchpole CK, Slater PJB. Bird song: Biological themes and variations. Second. Cambridge, U.K: Cambridge University Press; 2008.
-
-
Riebel K, Smallegange IM, Terpstra NJ, Bolhuis JJ. Sexual equality in zebra finch song preference: evidence for a dissociation between song recognition and production learning. Proceedings of the Royal Society B: Biological Sciences. 2002;269:729–733. Female Zebra Finches do not sing but choose to hear playback of their father's song in an operant task. The results show that fine auditory discrimination is learned and uncoupled from motor learning to sing.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
