

Review
doi: 10.1007/s10815-016-0652-1.
Epub 2016 Jan 29.
The axoneme: the propulsive engine of spermatozoa and cilia and associated ciliopathies leading to infertility
Affiliations
- PMID: 26825807
- PMCID: PMC4759005
- DOI: 10.1007/s10815-016-0652-1
Item in Clipboard
Review
The axoneme: the propulsive engine of spermatozoa and cilia and associated ciliopathies leading to infertility
J Assist Reprod Genet.
2016 Feb.
Abstract
This review article provides a critical analysis of the structure and molecular mechanisms of the microtubule axoneme of cilia and sperm flagella and their associated elements required for male fertility.A broad range of genetic and molecular defects (ciliopathies) are considered in the context of human diseases involving impaired motility in cilia and sperm flagella, providing provocative thought for future research in the area of male infertility.
Keywords: Basal body; Fibrous sheath; Intraflagellar transport; Microtubules; Outer dense fibers; Primary cilia.
Figures
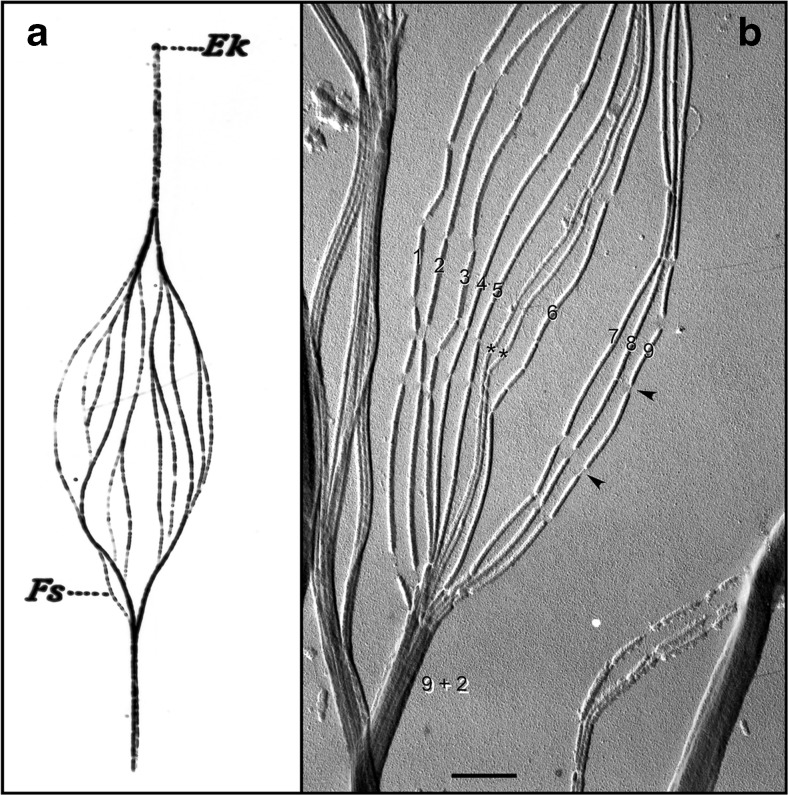
a Hand-drawn, light microscopic observations of a splayed rooster sperm flagellum by Ballowitz in 1888 [6], probably the first microscopic examination of sperm flagellar structure, showing the flagellum to be composed of 11 longitudinal elements (Fs). Ek, Endkörperchen (little end body) probably refers to what is now known as the basal body. b Electron micrograph (probably the first) of a splayed rooster sperm flagellum, by Grigg and Hodge in 1949 [7], showing 11 longitudinal elements, nine of which were noted to be wider/denser (numbered) than the other two (asterisks). This appearance suggested to the authors that the nine denser elements surrounded the two central less dense elements, also observed and diagrammed by Manton and Clark [8]—i.e., forming what would later be shown by Fawcett and Porter [9] to be the “9 + 2” microtubule axoneme. Grigg suggested to us that the thread-like connections (arrowheads) where the microtubule breaksdown (following protease treatment) might correspond to tektin filaments. Scale bar (b), 0.5 μm. Image courtesy of G.W. Grigg in 1998
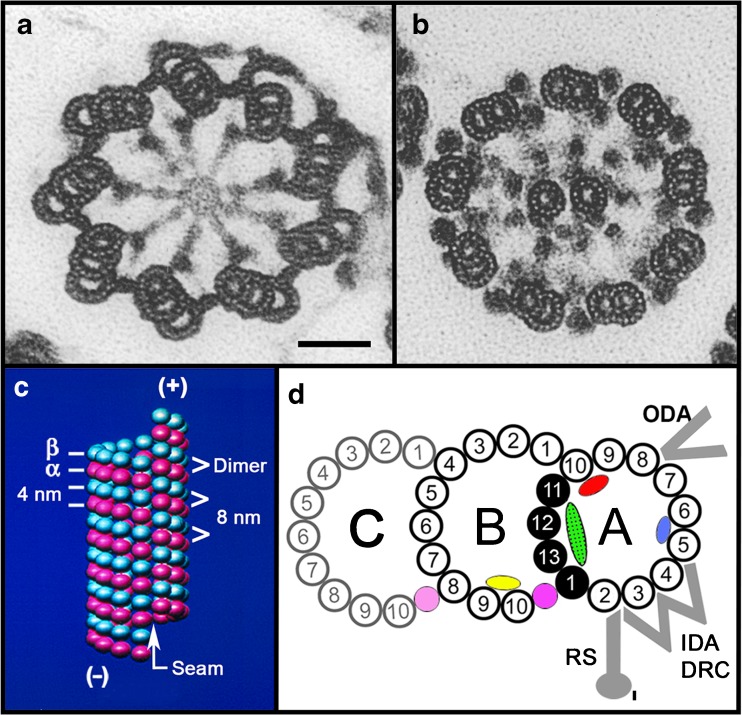
Structure of the basal body (a), the flagellar axoneme (b), a singlet microtubule (c), and a doublet microtubule (d). a, b Electron micrographs of cross sections of tannic acid-stained specimens (an Oxymonad protozoan flagellum) showing the ninefold arrangement of triplet microtubules of a basal body, the 9 + 2 axoneme, and the protofilament substructure of the microtubule walls (compare with Fig. 3). Note: As viewed from the basal body to the flagellar tip, the triplet microtubules are tilted in a clockwise pinwheel pattern and the dynein arms point in a clockwise direction toward their adjacent doublet microtubule. Scale bar for a and b, 50 nm; images taken by D. Woodrum Hensley—see [14]. c Illustrated are the arrangement and spacings of the α-tubulin and β-tubulin subunits (rendered as spheres) forming the αβ-dimers, their axial repeats (interprotofilament spacing, 5 nm), the lattice or arrangement of subunits around the microtubule wall, the seam or discontinuity in the lattice, and the plus and minus ends of the microtubule. d Diagram of the structure of the doublet and triplet microtubule (basal body C-tubule shaded): protofilaments numbered according to convention [15]; the Sarkosyl-insoluble Ribbon of four protofilaments (black) [16]; connections of the B-tubule to the A-tubule and the C-tubule to the B-tubule, as determined in a and b; microtubule inner proteins include MIP1 (blue), MIP2 (red), MIP3 (yellow), inner A-B junctional protein (pink), and inner B-C junctional proteins (faint pink) [10, 11]; partition-associated material (green) [16]; and approximate positions of the outer dynein arms (ODA), inner dynein arms (IDA), dynein regulatory complex (DRC), and radial spokes (RS) [12, 13]
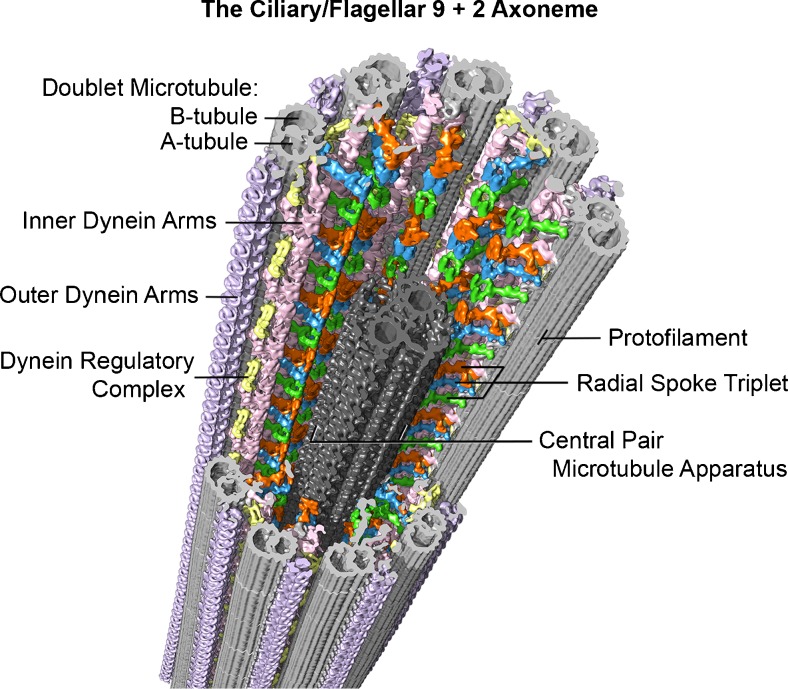
Example of the current, advanced imaging of the 9 + 2 axoneme (from sea urchin, Strongylocentrotus purpuratus, sperm flagella), using cryo-electron tomography with a resolution of approximately 3 nm. In this method, isolated flagella or axonemes, applied to special EM grids, are frozen within a few milliseconds in liquid ethane, which prevents damaging ice crystal formation. The specimen is then transferred to a cryo-transfer holder cooled with liquid nitrogen and inserted into the transmission electron microscope. After locating a promising area of a frozen flagellum or axoneme at medium magnification, a tilt series with up to 100 tilted views (from −65° to +65°) is recorded at higher magnification with low electron doses to minimize specimen radiation damage. The tilt series are then computationally aligned and the 3D structure of the specimen is reconstructed. The 96-nm longitudinal repeats of the axoneme (see text) are then extracted and averaged to increase the signal to noise ratio and thus resolution. Finally, the averaged repeat is visualized in 3D using isosurface rendering, as shown here. Some of the major structural features are labeled: Doublet A- and B-tubules (gray), radial spokes 1–3 (green, blue, orange), outer dynein arms (lavender), inner dynein arms (pink), nexin-dynein regulatory complex (yellow), and the central pair microtubule apparatus (charcoal). Image courtesy of Daniel Stoddard and Dr. Jianfeng Lin from the laboratory of Dr. Daniela Nicastro (Brandeis University and University of Texas Southwestern Medical Center). See references [–13]
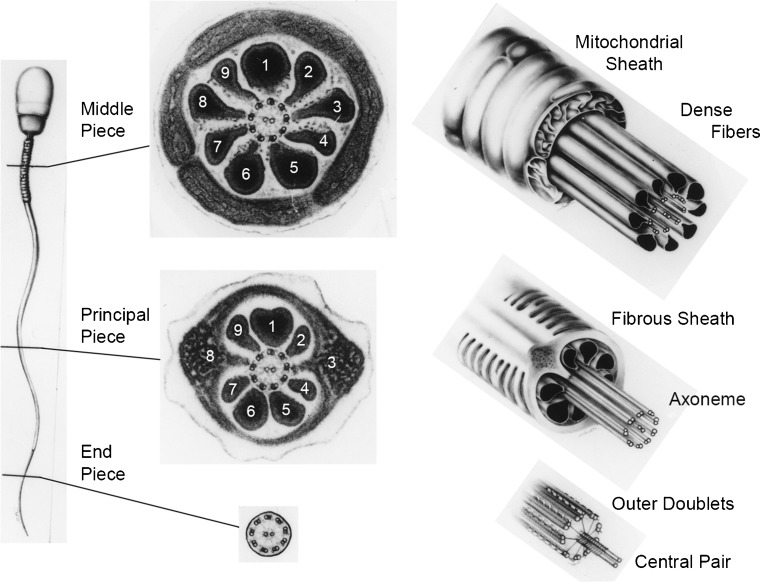
Electron micrographs of cross sections and diagrammatic 3D views of the mammalian (human) spermatozoon, showing the features and transitions occurring along the flagellum. Not illustrated is the fact that the outer dense fibers (ODFs) and the fibrous sheath (FS) gradually taper and terminate where the endpiece extends. Because of the unique morphologies of the ODFs, they and the outer doublet microtubules (to which the ODFs are attached) can be unequivocally numbered. Note also that doublet microtubules #3 and #8 are firmly anchored to their respective longitudinal columns of the FS. Courtesy of Dr. D.W. Fawcett. See references [27, 28]
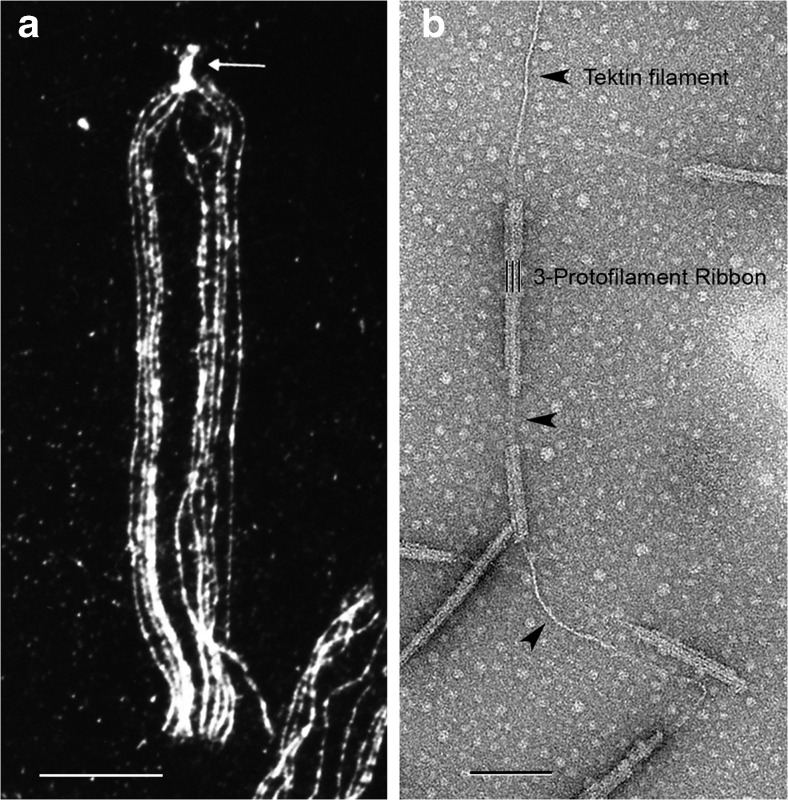
a Immunofluorescence light micrograph of a splayed sea urchin sperm flagellum (L. pictus) stained with anti-tektin-B antibody, showing the presence of tektin along all nine doublet microtubules (where the punctate staining is due to the masking of tektin epitopes by fixed tubulin) and a greater intensity of tektin staining in the basal body (arrow); image taken by W. Steffen [54]. b Electron micrograph of a partially fractionated Ribbon of three protofilaments (three black lines) of a sea urchin (S. purpuratus) sperm flagellar doublet microtubule, negatively stained, showing the single tektin filament (arrowheads) emerging from the end of the Ribbon and/or connecting segments of a disassembled Ribbon; image taken by A. Schefter in the laboratory of R.W. Linck—see [16]. The tektin filament may correspond to the thread-like connections in Fig. 1b. Scale bars 10 μm in a and 100 nm in b
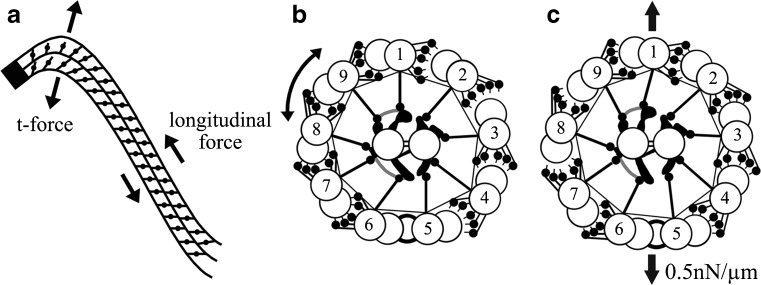
This figure, taken from Lindemann and Mitchell [92], illustrates several features and hypotheses of ciliary and flagellar bend formation and bend propagation. First in a, the nine doublet microtubules are anchored to the basal body (the solid black end), where they cannot slide initially, but the ability and degree with which they slide immediately increases as the distance from the basal body increases. Secondly, sliding takes place as dynein arms generate longitudinal sliding forces between doublet tubules, moving adjacent doublet tubules toward the base and generating a bend. Thirdly, because of the cylindrical arrangement of the axoneme, the interdoublet sliding will generate a transverse t-force that acts to compress or expand the diameter of axoneme. Finally in b and c, this t-force will alternately bring doublet tubules on one side of the axoneme closer together and permit dynein-tubule sliding, while on the other side, the doublet tubules are pushed slightly further apart, preventing dynein-tubule interactions; these two events then oscillate back and forth to generate propagated bends. These concepts are brought together in the Geometric Clutch hypothesis [93]. Courtesy of author C.B. Lindemann, and publisher, John Wiley & Sons, Ltd
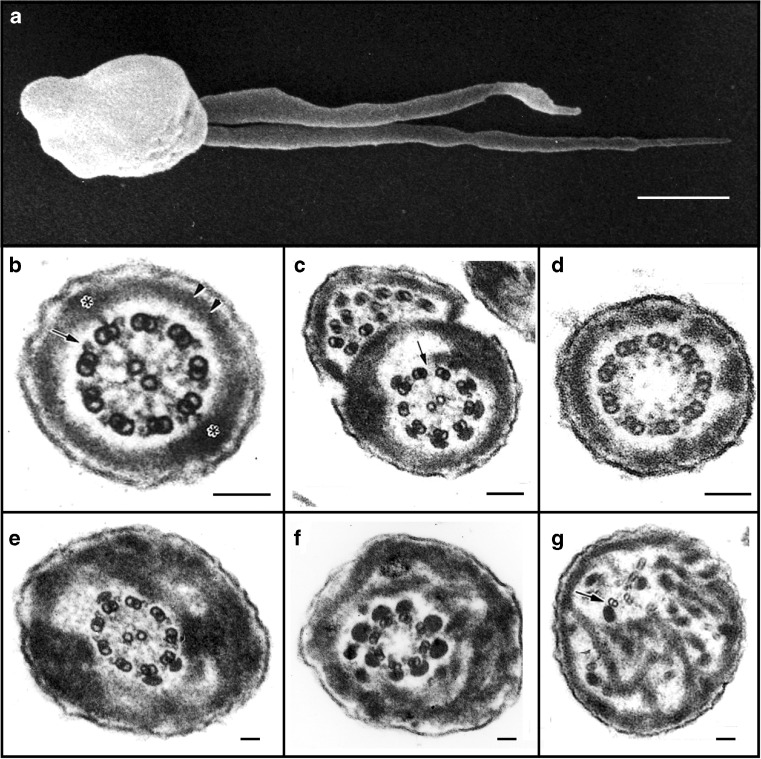
Electron microscopic analysis of human sperm pathologies. a Scanning electron micrograph of a dysplasia of the fibrous sheath (DFS) in human spermatozoa. Note the two thick, irregular and very short tails (length ˂10 μm, normal 50–60 μm). b Transverse section of a normal flagellum at the distal principal piece (compare with Fig. 4). At this point, the axoneme is composed of nine doublet microtubules around a central pair microtubule apparatus (CPMA), nine radial spokes projecting toward the CPMA, and two dynein arms (outer and inner) anchored to each A-tubule and oriented in a clockwise direction toward the B-tubule of the neighboring doublet microtubule (arrow), as viewed from the base to the tip of the flagellum. The FS is composed of two lateral columns inserting into doublet microtubules #3 and #8 (asterisks), and two semi-circumferential ribs (double arrow head). By this point along the flagellum, the ODFs have terminated. c, d Spermatozoa from two patients with primary ciliary dyskinesia. In c, the fused complete and incomplete axonemes are due to the failure of neighboring spermatids to separate from their cytoplasmic bridges, and here, there is partial lack of dynein arms (arrow) and FS distortions. In d, the CPMA is missing (i.e., a 9 + 0 axoneme) but radial spokes are still present. e–g Three transverse sections of DFS spermatozoa with marked FS abnormalities: the FS forms thick disordered periaxonemal rings and the lateral columns are misplaced in e and f; the axoneme in e is preserved, but in f, there is lack of one doublet microtubule and the CPMA is missing (8 + 0 axoneme). In g, note the complete disorientation of the axoneme, where one doublet microtubule appears to lack dynein arms (arrow). Diameters of pathological flagella may range from 1 to 1.2 μm (normal flagellar diameter ≅ 0.4 μm). Scale bars 1 μm in a and 0.1 μm in b–g
References
-
- van Leeuwenhoek, A. http://lensonleeuwenhoek.net/.
-
- Gray J. The movement of sea-urchin spermatozoa. J Exp Biol. 1955;32:775–801.
-
- Vogl AW, Vaid KS, Guttman JA. The Sertoli cell cytoskeleton. Adv Exp Med Biol. 2008;636:186–211. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
