

T Cell Receptor-induced Nuclear Factor κB (NF-κB) Signaling and Transcriptional Activation Are Regulated by STIM1- and Orai1-mediated Calcium Entry
- PMID: 26826124
- PMCID: PMC4861418
- DOI: 10.1074/jbc.M115.713008
T Cell Receptor-induced Nuclear Factor κB (NF-κB) Signaling and Transcriptional Activation Are Regulated by STIM1- and Orai1-mediated Calcium Entry
Abstract
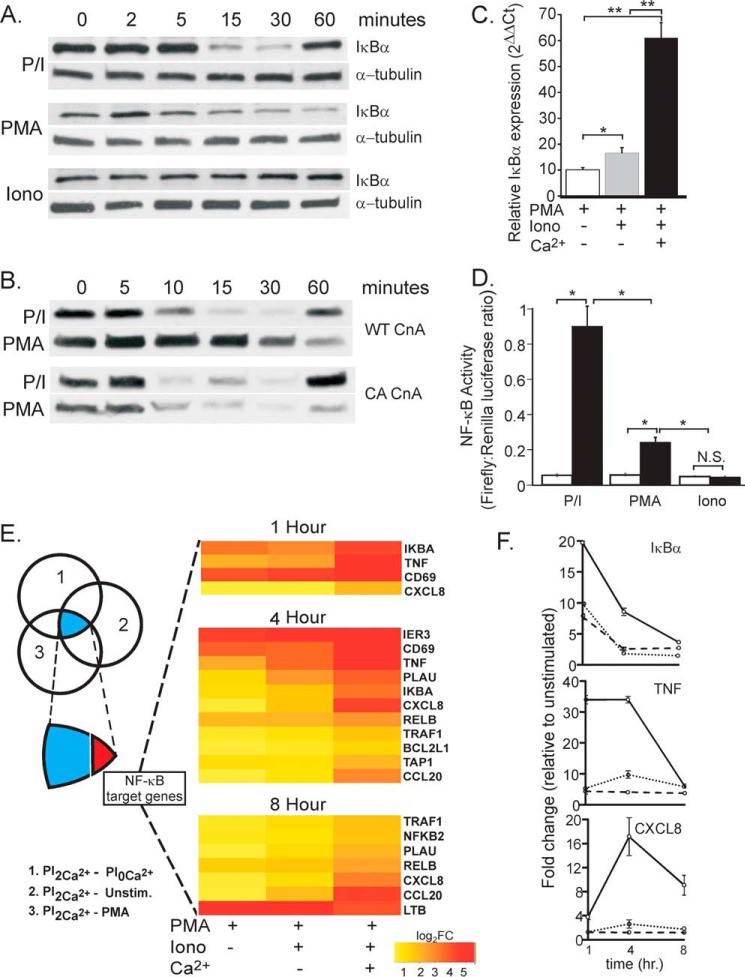
T cell activation following antigen binding to the T cell receptor (TCR) involves the mobilization of intracellular Ca(2+) to activate the key transcription factors nuclear factor of activated T lymphocytes (NFAT) and NF-κB. The mechanism of NFAT activation by Ca(2+) has been determined. However, the role of Ca(2+) in controlling NF-κB signaling is poorly understood, and the source of Ca(2+) required for NF-κB activation is unknown. We demonstrate that TCR- but not TNF-induced NF-κB signaling upstream of IκB kinase activation absolutely requires the influx of extracellular Ca(2+) via STIM1-dependent Ca(2+) release-activated Ca(2+)/Orai channels. We further show that Ca(2+) influx controls phosphorylation of the NF-κB protein p65 on Ser-536 and that this posttranslational modification controls its nuclear localization and transcriptional activation. Notably, our data reveal that this role for Ca(2+) is entirely separate from its upstream control of IκBα degradation, thereby identifying a novel Ca(2+)-dependent distal step in TCR-induced NF-κB activation. Finally, we demonstrate that this control of distal signaling occurs via Ca(2+)-dependent PKCα-mediated phosphorylation of p65. Thus, we establish the source of Ca(2+) required for TCR-induced NF-κB activation and define a new distal Ca(2+)-dependent checkpoint in TCR-induced NF-κB signaling that has broad implications for the control of immune cell development and T cell functional specificity.
Keywords: NF-κB transcription factor; PKC; T-cell receptor (TCR); calcium; calcium release-activated calcium channel protein 1 (ORAI1); posttranslational modification (PTM); stromal interaction molecule 1 (STIM1).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






Similar articles
-
Transcription factor NF-κB regulates expression of pore-forming Ca2+ channel unit, Orai1, and its activator, STIM1, to control Ca2+ entry and affect cellular functions.J Biol Chem. 2012 Jan 20;287(4):2719-30. doi: 10.1074/jbc.M111.275925. Epub 2011 Nov 21. J Biol Chem. 2012. PMID: 22110130 Free PMC article.
-
Analysis of Calcium Control of Canonical NF-κB Signaling in B Lymphocytes.Methods Mol Biol. 2021;2366:145-164. doi: 10.1007/978-1-0716-1669-7_9. Methods Mol Biol. 2021. PMID: 34236637
-
STIM- and Orai-mediated calcium entry controls NF-κB activity and function in lymphocytes.Cell Calcium. 2018 Sep;74:131-143. doi: 10.1016/j.ceca.2018.07.003. Epub 2018 Jul 10. Cell Calcium. 2018. PMID: 30048879 Free PMC article. Review.
-
Novel STIM1-dependent control of Ca2+ clearance regulates NFAT activity during T-cell activation.FASEB J. 2016 Nov;30(11):3878-3886. doi: 10.1096/fj.201600532R. Epub 2016 Aug 15. FASEB J. 2016. PMID: 27528628 Free PMC article.
-
ORAI-mediated calcium influx in T cell proliferation, apoptosis and tolerance.Cell Calcium. 2011 Sep;50(3):261-9. doi: 10.1016/j.ceca.2011.05.015. Epub 2011 Jun 12. Cell Calcium. 2011. PMID: 21665274 Review.
Cited by
-
The Blockade of Store-Operated Calcium Channels Improves Decompression Sickness in Rats.Front Physiol. 2020 Jan 31;10:1616. doi: 10.3389/fphys.2019.01616. eCollection 2019. Front Physiol. 2020. PMID: 32082179 Free PMC article.
-
4-1BB costimulation promotes CAR T cell survival through noncanonical NF-κB signaling.Sci Signal. 2020 Mar 31;13(625):eaay8248. doi: 10.1126/scisignal.aay8248. Sci Signal. 2020. PMID: 32234960 Free PMC article.
-
Calcium release-activated calcium channels and pain.Cell Calcium. 2018 Sep;74:180-185. doi: 10.1016/j.ceca.2018.07.009. Epub 2018 Jul 29. Cell Calcium. 2018. PMID: 30096536 Free PMC article. Review.
-
iCa2+ Flux, ROS and IL-10 Determines Cytotoxic, and Suppressor T Cell Functions in Chronic Human Viral Infections.Front Immunol. 2020 Mar 6;11:83. doi: 10.3389/fimmu.2020.00083. eCollection 2020. Front Immunol. 2020. PMID: 32210950 Free PMC article.
-
Transcriptomic dissection of termite gut microbiota following entomopathogenic fungal infection.Front Physiol. 2023 Apr 21;14:1194370. doi: 10.3389/fphys.2023.1194370. eCollection 2023. Front Physiol. 2023. PMID: 37153226 Free PMC article.
References
-
- Lewis R. S. (2003) Calcium oscillations in T-cells: mechanisms and consequences for gene expression. Biochem. Soc. Trans. 31, 925–929 - PubMed
-
- Healy J. I., Dolmetsch R. E., Lewis R. S., and Goodnow C. C. (1998) Quantitative and qualitative control of antigen receptor signalling in tolerant B lymphocytes. Novartis Found. Symp. 215, 137–144, discussion 144–145, 186–190 - PubMed
-
- Dolmetsch R. E., Lewis R. S., Goodnow C. C., and Healy J. I. (1997) Differential activation of transcription factors induced by Ca2+ response amplitude and duration. Nature 386, 855–858 - PubMed
-
- Healy J. I., Dolmetsch R. E., Timmerman L. A., Cyster J. G., Thomas M. L., Crabtree G. R., Lewis R. S., and Goodnow C. C. (1997) Different nuclear signals are activated by the B cell receptor during positive versus negative signaling. Immunity 6, 419–428 - PubMed
-
- Dolmetsch R. E., Xu K., and Lewis R. S. (1998) Calcium oscillations increase the efficiency and specificity of gene expression. Nature 392, 933–936 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
