

Searching for cognitive enhancement in the Morris water maze: better and worse performance in D-amino acid oxidase knockout (Dao(-/-)) mice
- PMID: 26833794
- PMCID: PMC4855640
- DOI: 10.1111/ejn.13192
Searching for cognitive enhancement in the Morris water maze: better and worse performance in D-amino acid oxidase knockout (Dao(-/-)) mice
Abstract
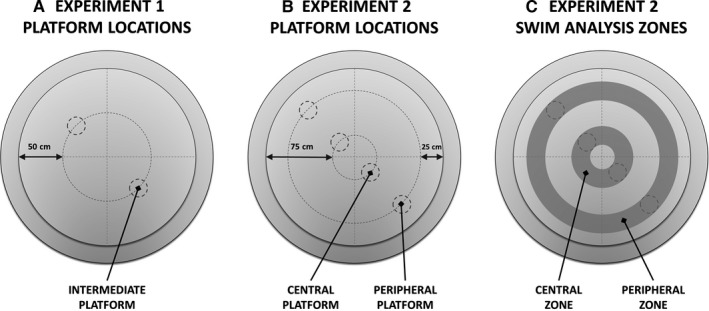
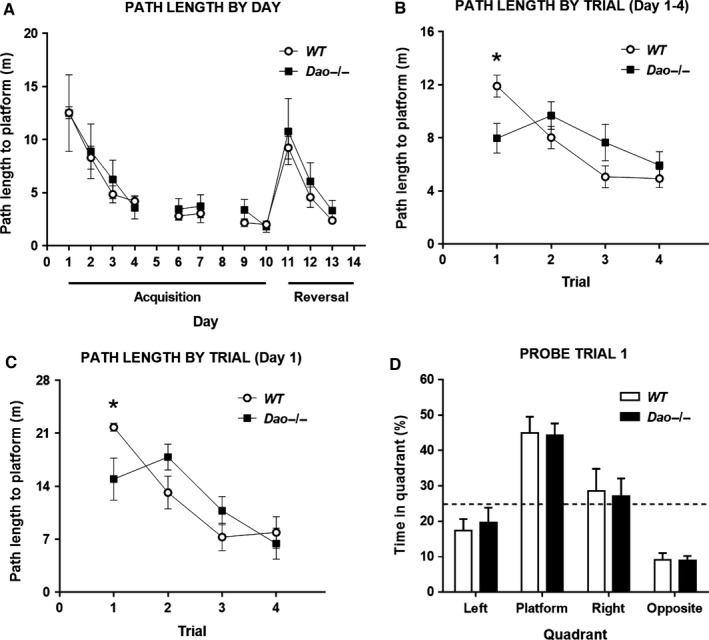
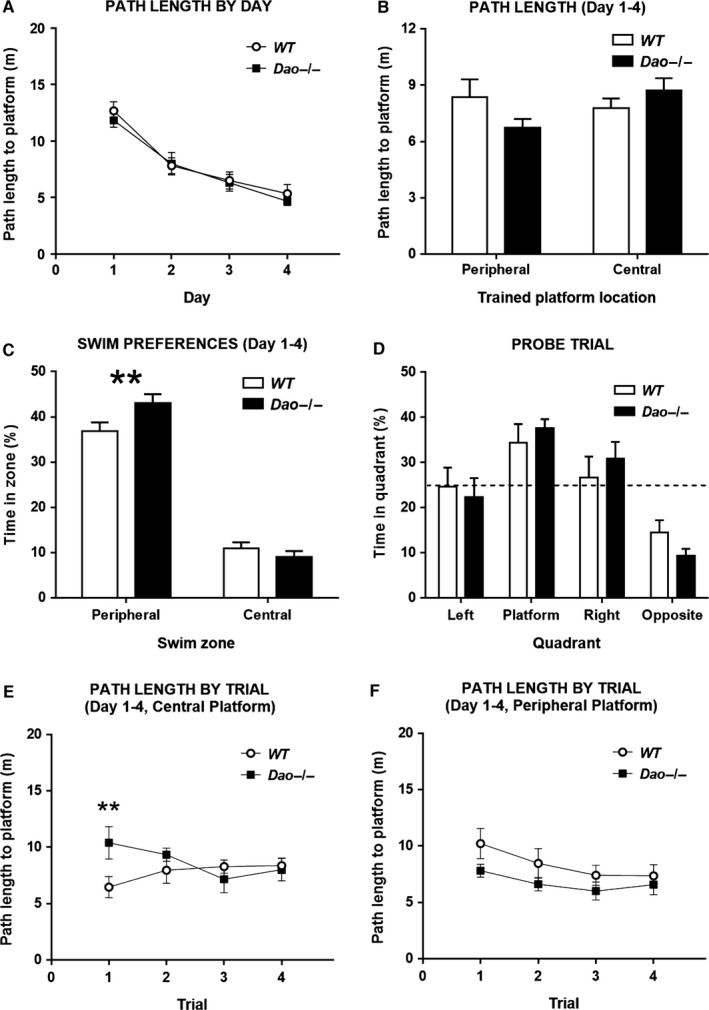
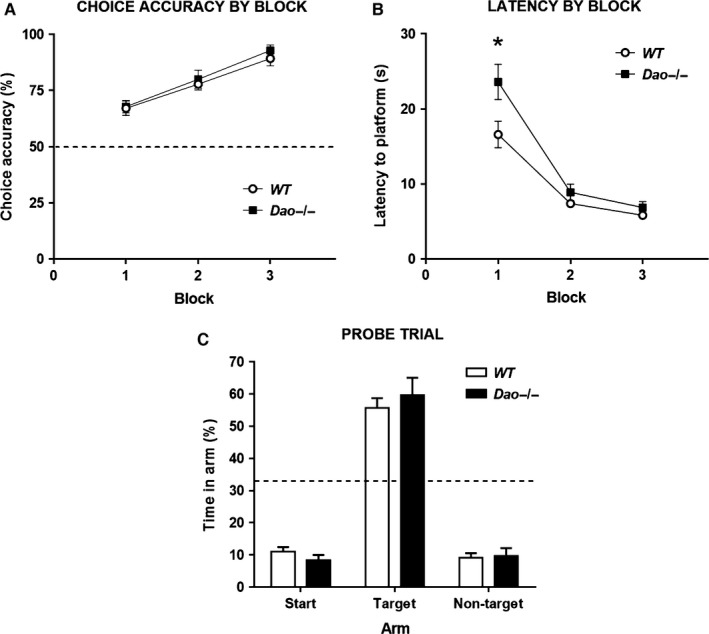
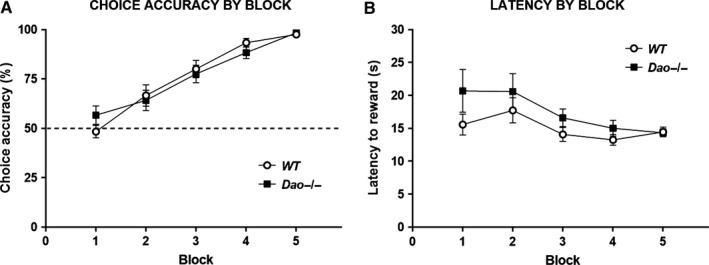
A common strategy when searching for cognitive-enhancing drugs has been to target the N-methyl-d-aspartate receptor (NMDAR), given its putative role in synaptic plasticity and learning. Evidence in favour of this approach has come primarily from studies with rodents using behavioural assays like the Morris water maze. D-amino acid oxidase (DAO) degrades neutral D-amino acids such as D-serine, the primary endogenous co-agonist acting at the glycine site of the synaptic NMDAR. Inhibiting DAO could therefore provide an effective and viable means of enhancing cognition, particularly in disorders like schizophrenia, in which NMDAR hypofunction is implicated. Indirect support for this notion comes from the enhanced hippocampal long-term potentiation and facilitated water maze acquisition of ddY/Dao(-) mice, which lack DAO activity due to a point mutation in the gene. Here, in Dao knockout (Dao(-/-) ) mice, we report both better and worse water maze performance, depending on the radial distance of the hidden platform from the side wall of the pool. Dao(-/-) mice displayed an increased innate preference for swimming in the periphery of the maze (possibly due to heightened anxiety), which facilitated the discovery of a peripherally located platform, but delayed the discovery of a centrally located platform. By contrast, Dao(-/-) mice exhibited normal performance in two alternative assays of long-term spatial memory: the appetitive and aversive Y-maze reference memory tasks. Taken together, these results question the proposed relationship between DAO inactivation and enhanced long-term associative spatial memory. They also have generic implications for how Morris water maze studies are performed and interpreted.
Keywords: D-serine; N-methyl-d-aspartate receptor; glutamate; pre-clinical model; schizophrenia.
© 2016 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures

Similar articles
-
d-amino acid oxidase knockout (Dao(-/-) ) mice show enhanced short-term memory performance and heightened anxiety, but no sleep or circadian rhythm disruption.Eur J Neurosci. 2015 May;41(9):1167-79. doi: 10.1111/ejn.12880. Epub 2015 Mar 27. Eur J Neurosci. 2015. PMID: 25816902 Free PMC article.
-
Genetic inactivation of D-amino acid oxidase enhances extinction and reversal learning in mice.Learn Mem. 2008 Dec 30;16(1):28-37. doi: 10.1101/lm.1112209. Print 2009 Jan. Learn Mem. 2008. PMID: 19117914 Free PMC article.
-
Mutant mice with reduced NMDA-NR1 glycine affinity or lack of D-amino acid oxidase function exhibit altered anxiety-like behaviors.Pharmacol Biochem Behav. 2009 Feb;91(4):610-20. doi: 10.1016/j.pbb.2008.09.016. Epub 2008 Oct 7. Pharmacol Biochem Behav. 2009. PMID: 18940194
-
D-Amino acids in the brain and mutant rodents lacking D-amino-acid oxidase activity.Amino Acids. 2012 Nov;43(5):1811-21. doi: 10.1007/s00726-012-1384-x. Epub 2012 Aug 15. Amino Acids. 2012. PMID: 22892863 Review.
-
The neurobiology of D-amino acid oxidase and its involvement in schizophrenia.Mol Psychiatry. 2010 Feb;15(2):122-37. doi: 10.1038/mp.2009.99. Epub 2009 Sep 29. Mol Psychiatry. 2010. PMID: 19786963 Free PMC article. Review.
Cited by
-
NMDARs, Coincidence Detectors of Astrocytic and Neuronal Activities.Int J Mol Sci. 2021 Jul 6;22(14):7258. doi: 10.3390/ijms22147258. Int J Mol Sci. 2021. PMID: 34298875 Free PMC article. Review.
-
Behavioral Analysis of NMDAR Function in Rodents: Tests of Long-Term Spatial Memory.Methods Mol Biol. 2024;2799:107-138. doi: 10.1007/978-1-0716-3830-9_7. Methods Mol Biol. 2024. PMID: 38727905
-
Transferrin Receptor 2 Dependent Alterations of Brain Iron Metabolism Affect Anxiety Circuits in the Mouse.Sci Rep. 2016 Aug 1;6:30725. doi: 10.1038/srep30725. Sci Rep. 2016. PMID: 27477597 Free PMC article.
-
Lack of the peroxiredoxin 6 gene causes impaired spatial memory and abnormal synaptic plasticity.Mol Brain. 2021 Apr 19;14(1):72. doi: 10.1186/s13041-021-00779-6. Mol Brain. 2021. PMID: 33874992 Free PMC article.
-
Improvement of cognitive deficit of curcumin on scopolamine-induced Alzheimer's disease models.Caspian J Intern Med. 2022 Winter;13(1):16-22. doi: 10.22088/cjim.13.1.16. Caspian J Intern Med. 2022. PMID: 35178203 Free PMC article.
References
-
- Bado, P. , Madeira, C. , Vargas‐Lopes, C. , Moulin, T.C. , Wasilewska‐Sampaio, A.P. , Maretti, L. , de Oliveira, R.V. , Amaral, O.B. & Panizzutti, R. (2011) Effects of low‐dose D‐serine on recognition and working memory in mice. Psychopharmacology, 218, 461–470. - PubMed
-
- Bannerman, D.M. , Rawlins, J.N. & Good, M.A. (2006) The drugs don't work ‐ or do they? Pharmacological and transgenic studies of the contribution of NMDA and GluR‐A‐containing AMPA receptors to hippocampal‐dependent memory. Psychopharmacology, 188, 552–566. - PubMed
-
- Bannerman, D.M. , Niewoehner, B. , Lyon, L. , Romberg, C. , Schmitt, W.B. , Taylor, A. , Sanderson, D.J. , Cottam, J. , Sprengel, R. , Seeburg, P.H. , Kohr, G. & Rawlins, J.N. (2008) NMDA receptor subunit NR2A is required for rapidly acquired spatial working memory but not incremental spatial reference memory. J. Neurosci., 28, 3623–3630. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
