

Functional and molecular characterization of mouse Gata2-independent hematopoietic progenitors
- PMID: 26834239
- PMCID: PMC4797020
- DOI: 10.1182/blood-2015-10-673749
Functional and molecular characterization of mouse Gata2-independent hematopoietic progenitors
Abstract
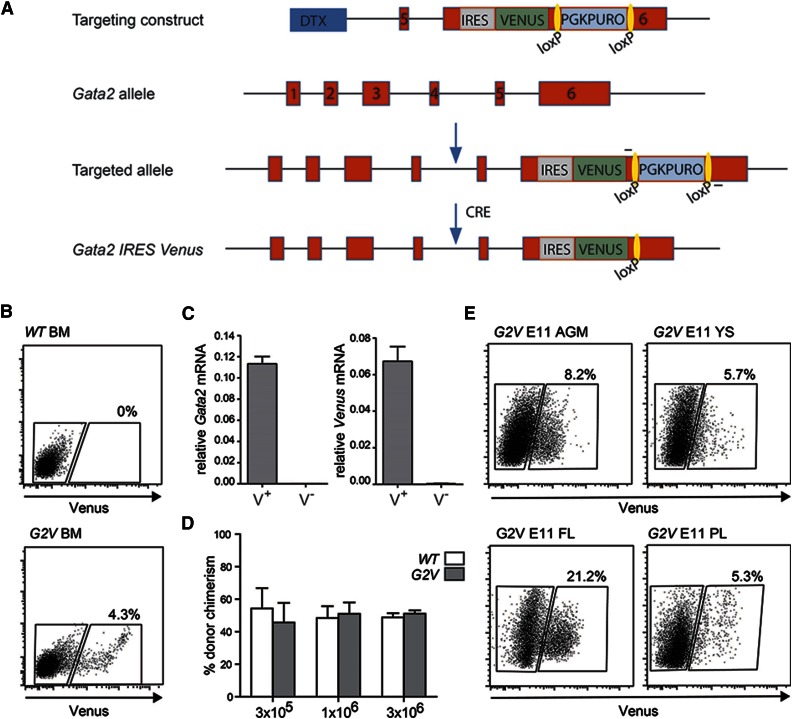
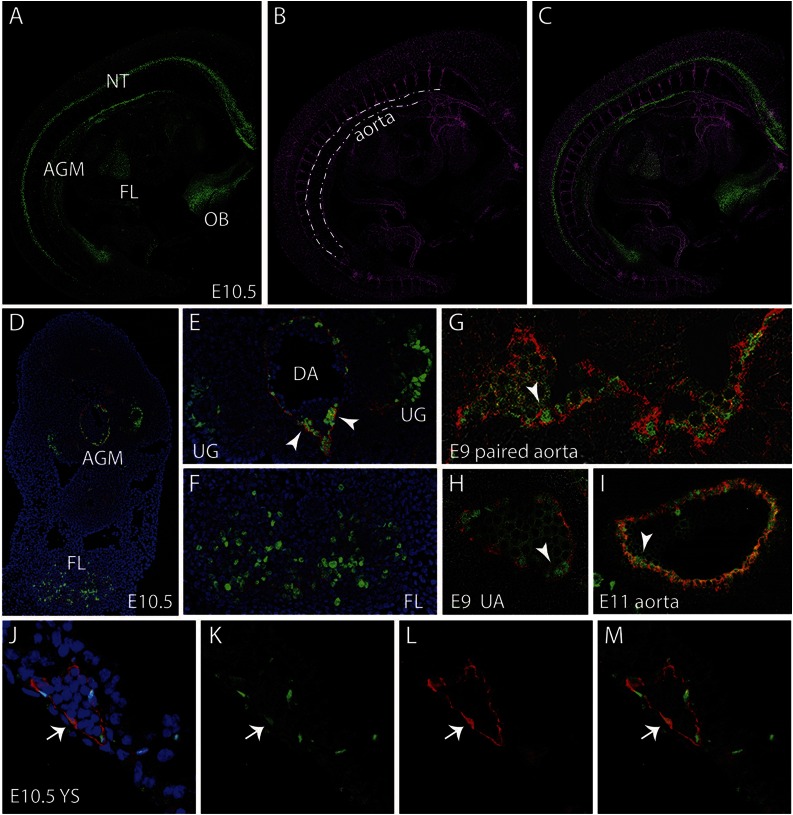
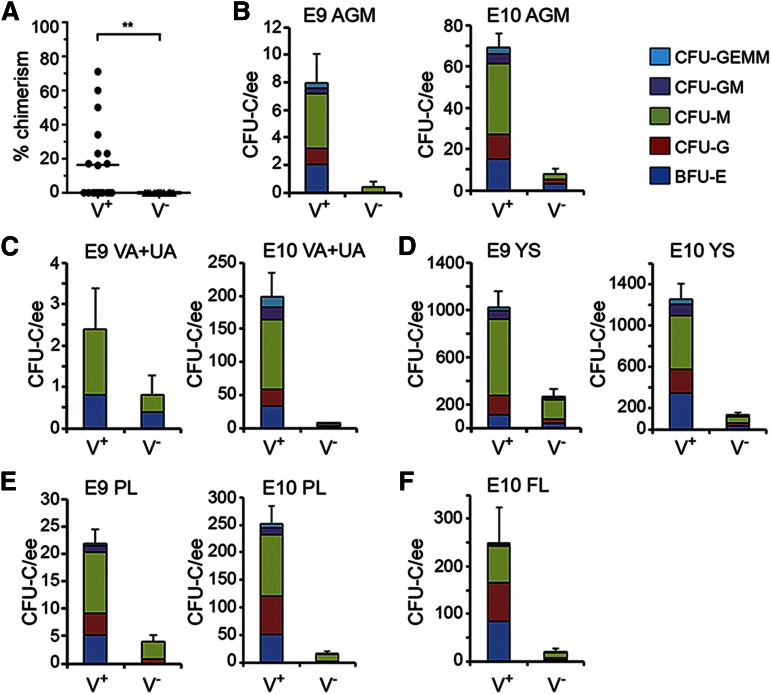
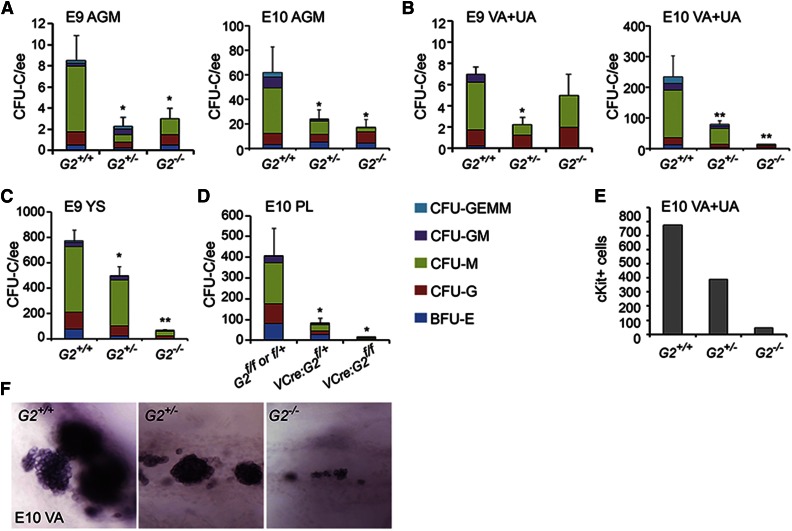
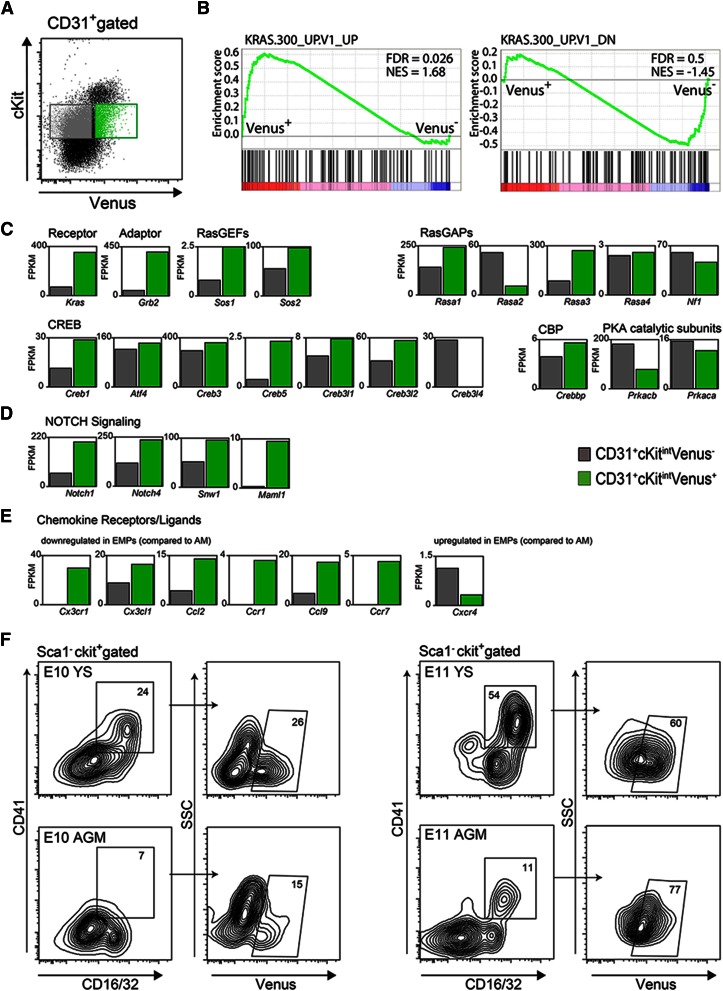
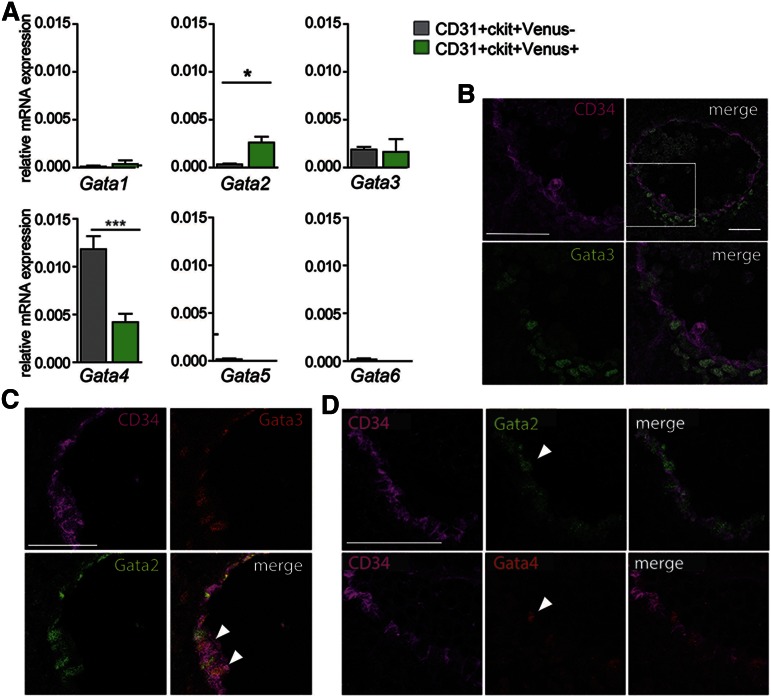
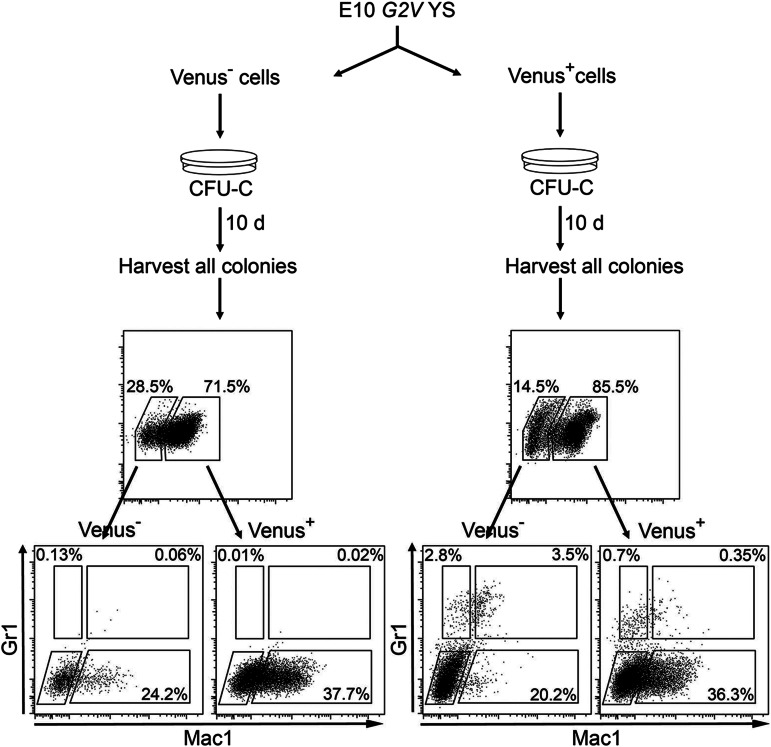
The Gata2 transcription factor is a pivotal regulator of hematopoietic cell development and maintenance, highlighted by the fact that Gata2 haploinsufficiency has been identified as the cause of some familial cases of acute myelogenous leukemia/myelodysplastic syndrome and in MonoMac syndrome. Genetic deletion in mice has shown that Gata2 is pivotal to the embryonic generation of hematopoietic stem cells (HSCs) and hematopoietic progenitor cells (HPCs). It functions in the embryo during endothelial cell to hematopoietic cell transition to affect hematopoietic cluster, HPC, and HSC formation. Gata2 conditional deletion and overexpression studies show the importance of Gata2 levels in hematopoiesis, during all developmental stages. Although previous studies of cell populations phenotypically enriched in HPCs and HSCs show expression of Gata2, there has been no direct study of Gata2 expressing cells during normal hematopoiesis. In this study, we generate a Gata2Venus reporter mouse model with unperturbed Gata2 expression to examine the hematopoietic function and transcriptome of Gata2 expressing and nonexpressing cells. We show that all the HSCs are Gata2 expressing. However, not all HPCs in the aorta, vitelline and umbilical arteries, and fetal liver require or express Gata2. These Gata2-independent HPCs exhibit a different functional output and genetic program, including Ras and cyclic AMP response element-binding protein pathways and other Gata factors, compared with Gata2-dependent HPCs. Our results, indicating that Gata2 is of major importance in programming toward HSC fate but not in all cells with HPC fate, have implications for current reprogramming strategies.
© 2016 by The American Society of Hematology.
Figures
Similar articles
-
Gata2 is required for HSC generation and survival.J Exp Med. 2013 Dec 16;210(13):2843-50. doi: 10.1084/jem.20130751. Epub 2013 Dec 2. J Exp Med. 2013. PMID: 24297996 Free PMC article.
-
Cis-element mutated in GATA2-dependent immunodeficiency governs hematopoiesis and vascular integrity.J Clin Invest. 2012 Oct;122(10):3692-704. doi: 10.1172/JCI61623. Epub 2012 Sep 10. J Clin Invest. 2012. PMID: 22996659 Free PMC article.
-
Conditional Gata2 inactivation results in HSC loss and lymphatic mispatterning.J Clin Invest. 2012 Oct;122(10):3705-17. doi: 10.1172/JCI61619. Epub 2012 Sep 10. J Clin Invest. 2012. PMID: 22996665 Free PMC article.
-
SCL/TAL1 in Hematopoiesis and Cellular Reprogramming.Curr Top Dev Biol. 2016;118:163-204. doi: 10.1016/bs.ctdb.2016.01.004. Epub 2016 Feb 18. Curr Top Dev Biol. 2016. PMID: 27137657 Review.
-
Developmental origins of hematopoietic stem cells.Oncol Res. 2003;13(6-10):315-21. doi: 10.3727/096504003108748519. Oncol Res. 2003. PMID: 12725520 Review.
Cited by
-
A Novel GATA2 Protein Reporter Mouse Reveals Hematopoietic Progenitor Cell Types.Stem Cell Reports. 2020 Aug 11;15(2):326-339. doi: 10.1016/j.stemcr.2020.06.008. Epub 2020 Jul 9. Stem Cell Reports. 2020. PMID: 32649900 Free PMC article.
-
The Il6 -39 kb enhancer containing clustered GATA2- and PU.1-binding sites is essential for Il6 expression in murine mast cells.iScience. 2022 Aug 14;25(9):104942. doi: 10.1016/j.isci.2022.104942. eCollection 2022 Sep 16. iScience. 2022. PMID: 36072552 Free PMC article.
-
Hematopoietic (stem) cell development - how divergent are the roads taken?FEBS Lett. 2016 Nov;590(22):3975-3986. doi: 10.1002/1873-3468.12372. Epub 2016 Sep 1. FEBS Lett. 2016. PMID: 27543859 Free PMC article. Review.
-
Autophagy regulates the maturation of hematopoietic precursors in the embryo.Nat Commun. 2024 Mar 15;15(1):2255. doi: 10.1038/s41467-024-46453-y. Nat Commun. 2024. PMID: 38490977 Free PMC article.
-
Hematological phenotypes in GATA2 deficiency syndrome arise from aging, maladaptation to proliferation, and somatic events.Blood Adv. 2025 Jun 10;9(11):2794-2807. doi: 10.1182/bloodadvances.2024015106. Blood Adv. 2025. PMID: 40138552 Free PMC article.
References
-
- Wilson NK, Foster SD, Wang X, et al. Combinatorial transcriptional control in blood stem/progenitor cells: genome-wide analysis of ten major transcriptional regulators. Cell Stem Cell. 2010;7(4):532–544. - PubMed
-
- Fujiwara Y, Chang AN, Williams AM, Orkin SH. Functional overlap of GATA-1 and GATA-2 in primitive hematopoietic development. Blood. 2004;103(2):583–585. - PubMed
-
- Tsai FY, Keller G, Kuo FC, et al. An early haematopoietic defect in mice lacking the transcription factor GATA-2. Nature. 1994;371(6494):221–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
