

Parallel Olfactory Processing in the Honey Bee Brain: Odor Learning and Generalization under Selective Lesion of a Projection Neuron Tract
- PMID: 26834589
- PMCID: PMC4717326
- DOI: 10.3389/fnint.2015.00075
Parallel Olfactory Processing in the Honey Bee Brain: Odor Learning and Generalization under Selective Lesion of a Projection Neuron Tract
Abstract
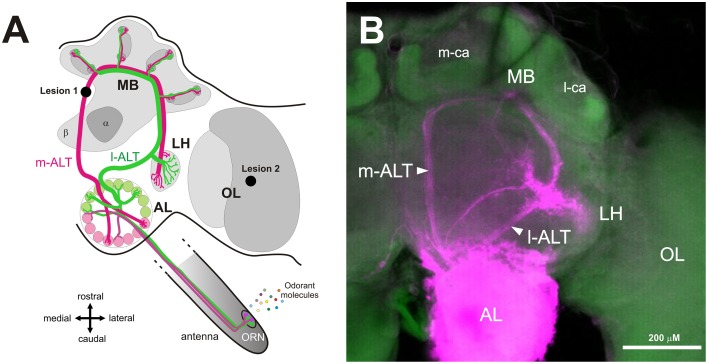
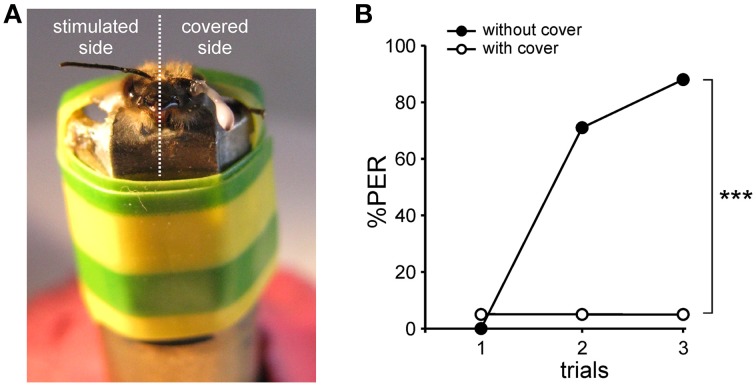
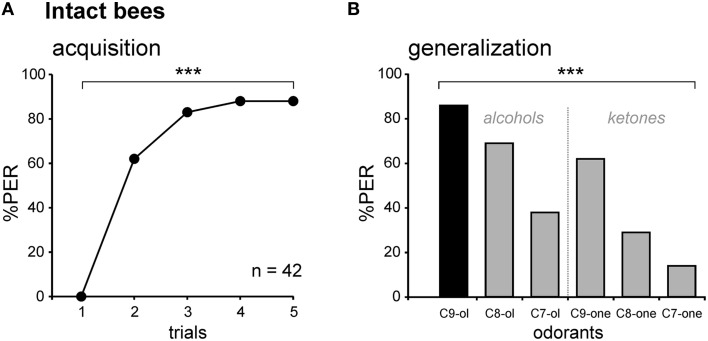
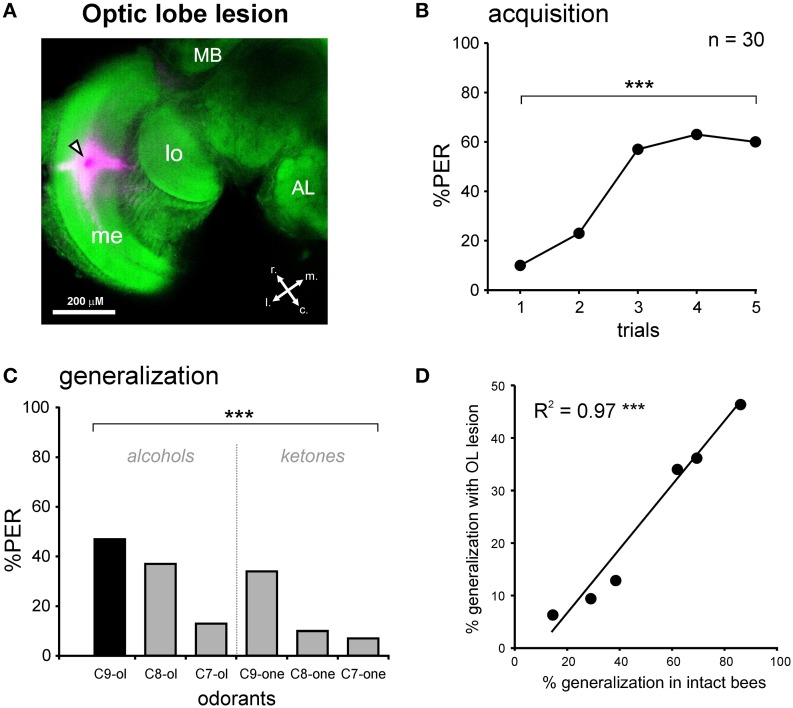
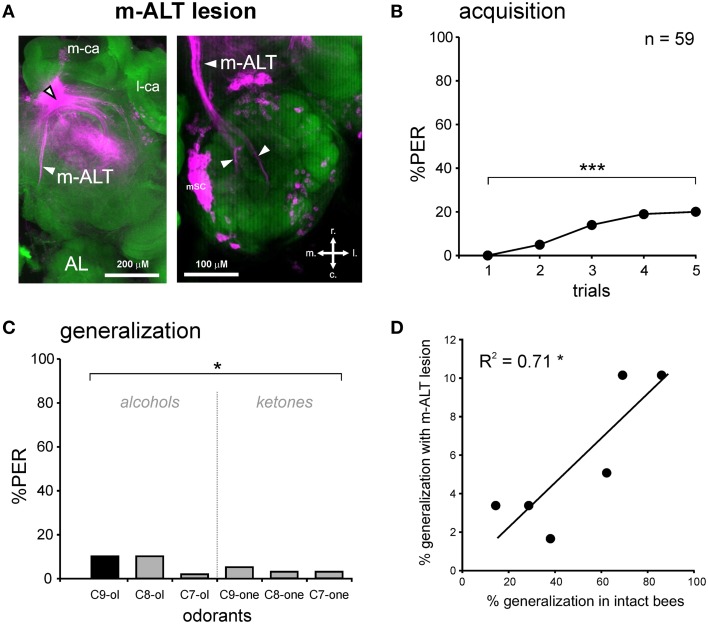
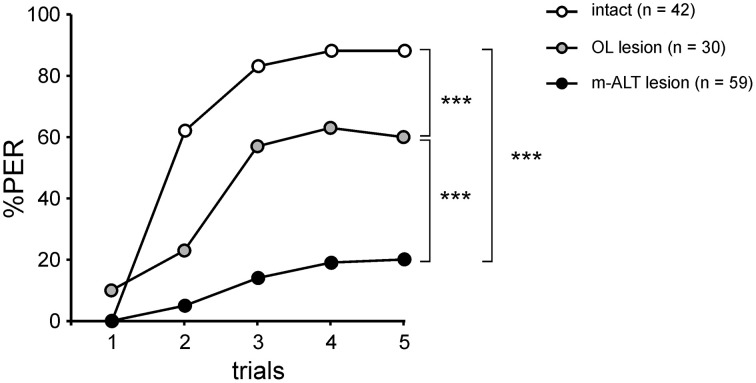
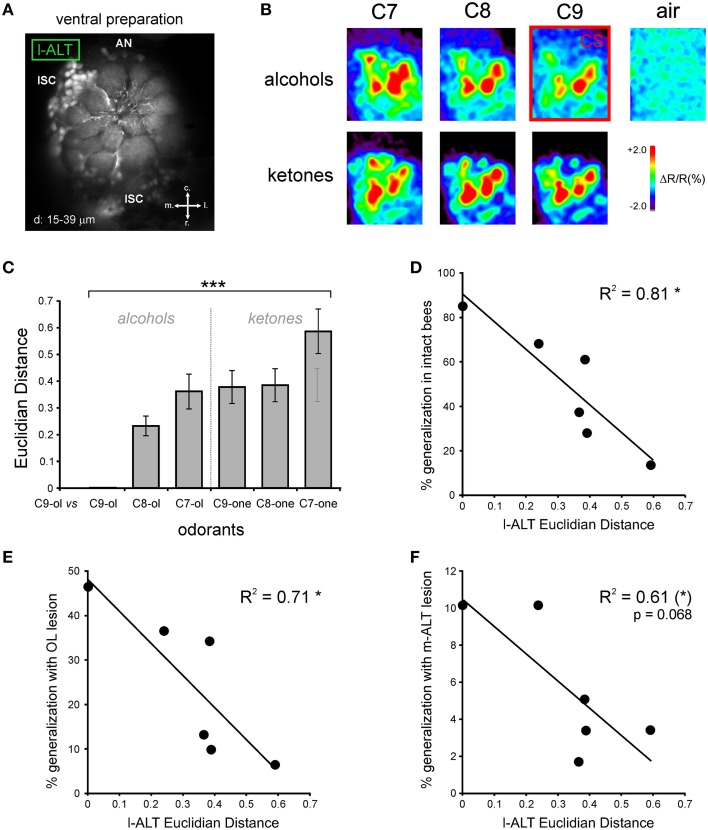
The function of parallel neural processing is a fundamental problem in Neuroscience, as it is found across sensory modalities and evolutionary lineages, from insects to humans. Recently, parallel processing has attracted increased attention in the olfactory domain, with the demonstration in both insects and mammals that different populations of second-order neurons encode and/or process odorant information differently. Among insects, Hymenoptera present a striking olfactory system with a clear neural dichotomy from the periphery to higher-order centers, based on two main tracts of second-order (projection) neurons: the medial and lateral antennal lobe tracts (m-ALT and l-ALT). To unravel the functional role of these two pathways, we combined specific lesions of the m-ALT tract with behavioral experiments, using the classical conditioning of the proboscis extension response (PER conditioning). Lesioned and intact bees had to learn to associate an odorant (1-nonanol) with sucrose. Then the bees were subjected to a generalization procedure with a range of odorants differing in terms of their carbon chain length or functional group. We show that m-ALT lesion strongly affects acquisition of an odor-sucrose association. However, lesioned bees that still learned the association showed a normal gradient of decreasing generalization responses to increasingly dissimilar odorants. Generalization responses could be predicted to some extent by in vivo calcium imaging recordings of l-ALT neurons. The m-ALT pathway therefore seems necessary for normal classical olfactory conditioning performance.
Keywords: insect; olfaction; olfactory conditioning; parallel processing; projection neurons; specific lesion.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources