

Ethylene, a Hormone at the Center-Stage of Nodulation
- PMID: 26834752
- PMCID: PMC4714629
- DOI: 10.3389/fpls.2015.01121
Ethylene, a Hormone at the Center-Stage of Nodulation
Abstract
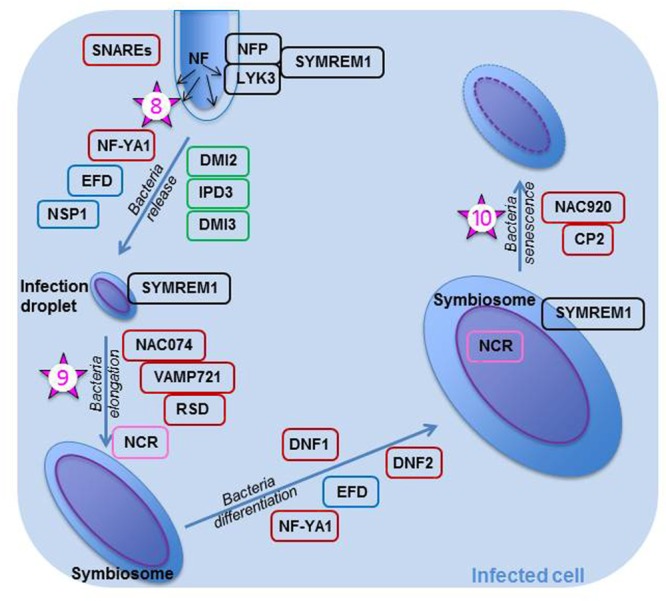
Nodulation is the result of a beneficial interaction between legumes and rhizobia. It is a sophisticated process leading to nutrient exchange between the two types of symbionts. In this association, within a nodule, the rhizobia, using energy provided as photosynthates, fix atmospheric nitrogen and convert it to ammonium which is available to the plant. Nodulation is recognized as an essential process in nitrogen cycling and legume crops are known to enrich agricultural soils in nitrogenous compounds. Furthermore, as they are rich in nitrogen, legumes are considered important as staple foods for humans and fodder for animals. To tightly control this association and keep it mutualistic, the plant uses several means, including hormones. The hormone ethylene has been known as a negative regulator of nodulation for almost four decades. Since then, much progress has been made in the understanding of both the ethylene signaling pathway and the nodulation process. Here I have taken a large view, using recently obtained knowledge, to describe in some detail the major stages of the process. I have not only reviewed the steps most commonly covered (the common signaling transduction pathway, and the epidermal and cortical programs), but I have also looked into steps less understood (the pre-infection step with the plant defense response, the bacterial release and the formation of the symbiosome, and nodule functioning and senescence). After a succinct review of the ethylene signaling pathway, I have used the knowledge obtained from nodulation- and ethylene-related mutants to paint a more complete picture of the role played by the hormone in nodule organogenesis, functioning, and senescence. It transpires that ethylene is at the center of this effective symbiosis. It has not only been involved in most of the steps leading to a mature nodule, but it has also been implicated in host immunity and nodule senescence. It is likely responsible for the activation of other hormonal signaling pathways. I have completed the review by citing three studies which makes one wonder whether knowledge gained on nodulation in the last decades is ready to be transferred to agricultural fields.
Keywords: ethylene signaling; hormones; host immunity; model legumes; nodule organogenesis; nodule senescence; rhizobia; sickle.
Figures


References
-
- Abeles F. B., Morgan P. W., Saltweit M. E. (1992). Ethylene in Plant Biology, 2nd Edn. San Diego, CA: Academic Press.
-
- Arora N. K., Khare E., Singh S., Maheshwari D. K. (2010). Effect of Al and heavy metals on enzymes of nitrogen metabolism of fast and slow-growing rhizobia under explanta conditions. World J. Microbiol. Microtechnol. 26 811–816. 10.1007/s11274-009-0237-6 - DOI
-
- Aziz M. Z., Saleem M., Ahmad Z., Yaseen M., Naveed M., Khan M. Y., et al. (2015). Effect of ethylene on growth, nodulation, early flower induction and yield in mungbean. Int. J. Sci. Eng. Res. 6 1210–1224.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources