

Proaggregant nuclear factor(s) trigger rapid formation of α-synuclein aggregates in apoptotic neurons
- PMID: 26839082
- PMCID: PMC4911378
- DOI: 10.1007/s00401-016-1542-4
Proaggregant nuclear factor(s) trigger rapid formation of α-synuclein aggregates in apoptotic neurons
Abstract
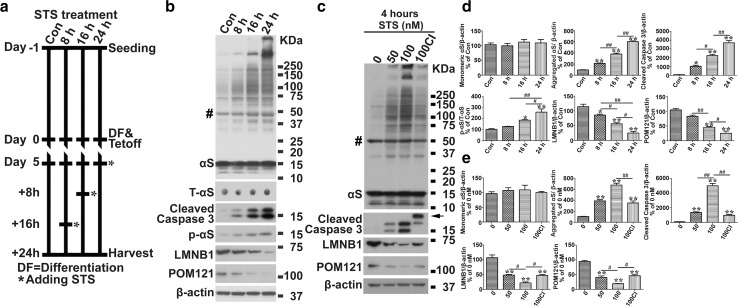
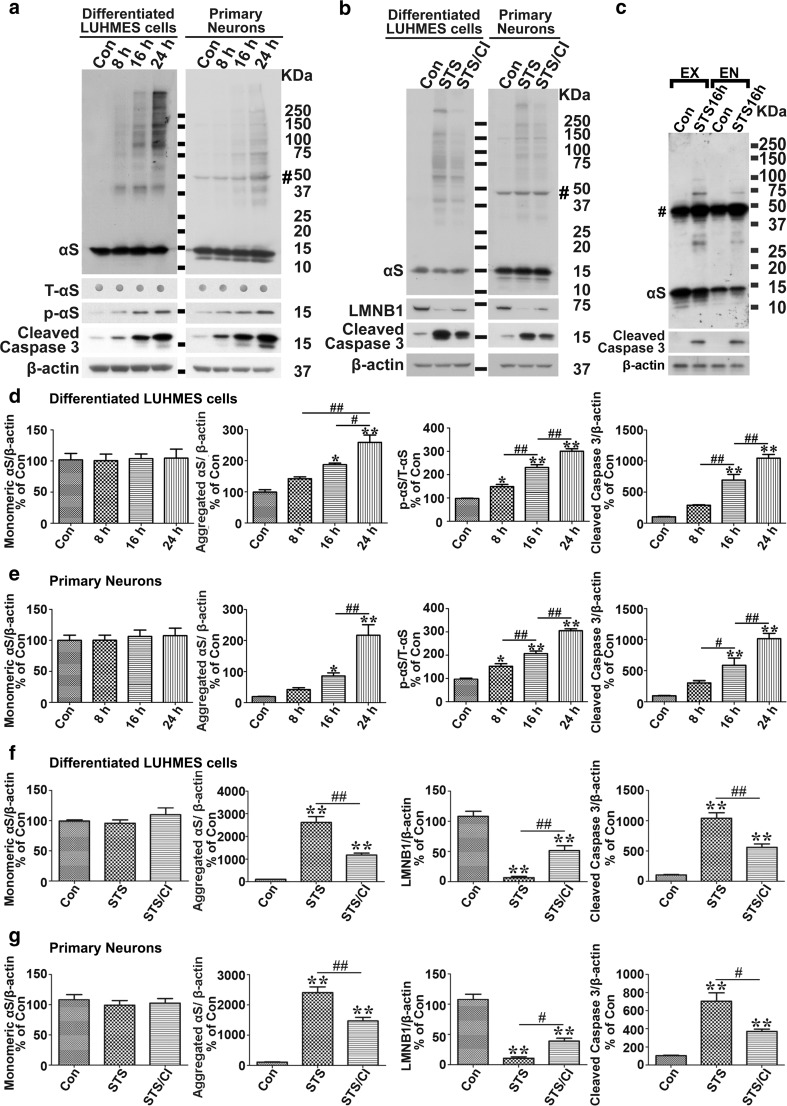
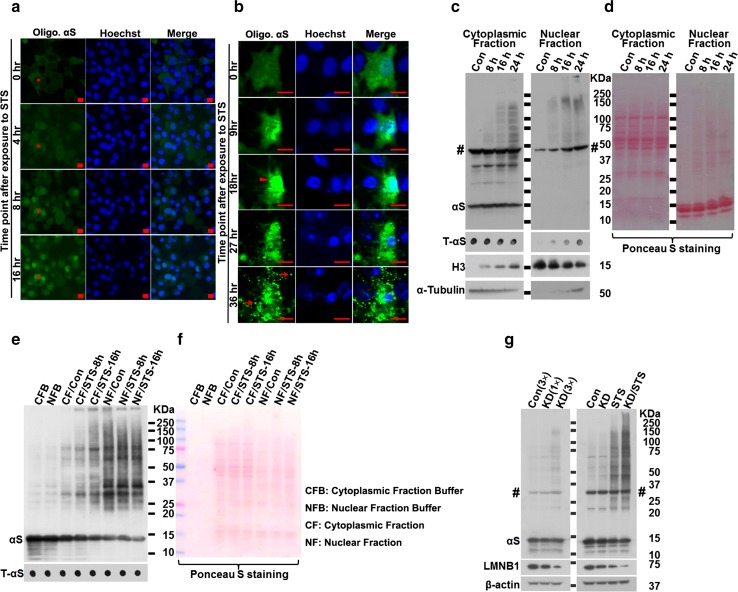
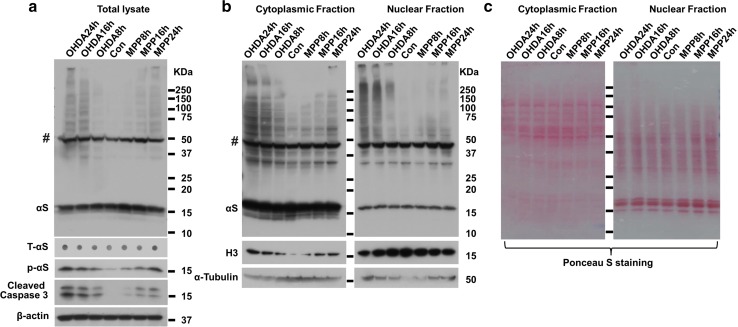
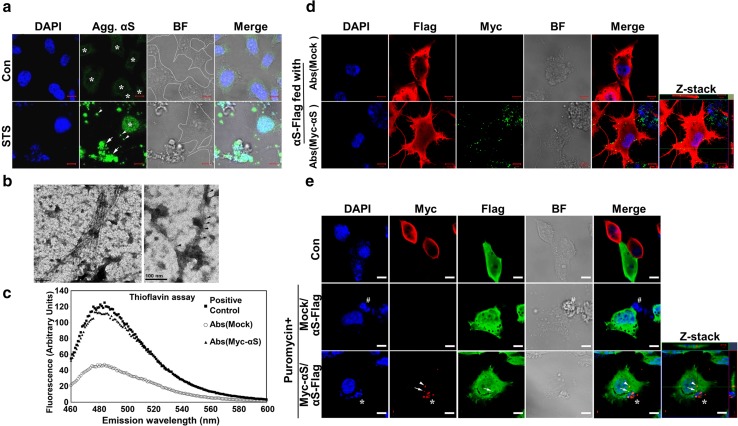
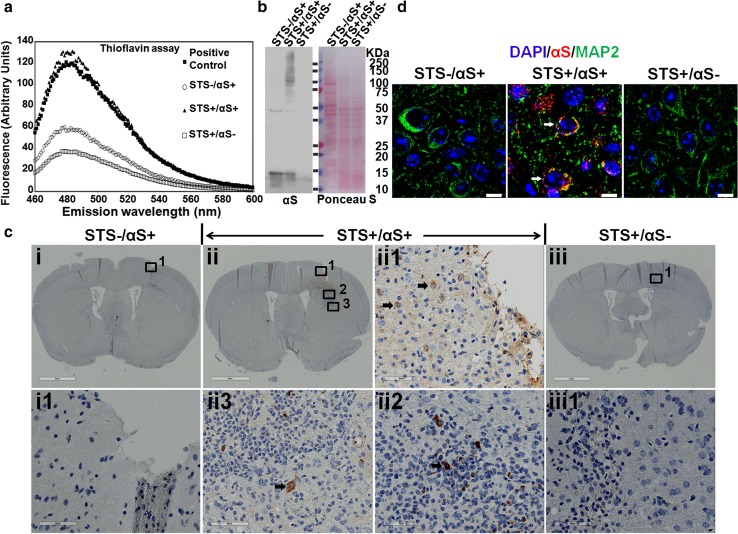
Cell-to-cell transmission of α-synuclein (αS) aggregates has been proposed to be responsible for progressive αS pathology in Parkinson disease (PD) and related disorders, including dementia with Lewy bodies. In support of this concept, a growing body of in vitro and in vivo experimental evidence shows that exogenously introduced αS aggregates can spread into surrounding cells and trigger PD-like pathology. It remains to be determined what factor(s) lead to initiation of αS aggregation that is capable of seeding subsequent propagation. In this study we demonstrate that filamentous αS aggregates form in neurons in response to apoptosis induced by staurosporine or other toxins-6-hydroxy-dopamine and 1-methyl-4-phenylpyridinium (MPP+). Interaction between αS and proaggregant nuclear factor(s) is associated with disruption of nuclear envelope integrity. Knocking down a key nuclear envelop constituent protein, lamin B1, enhances αS aggregation. Moreover, in vitro and in vivo experimental models demonstrate that aggregates released upon cell breakdown can be taken up by surrounding cells. Accordingly, we suggest that at least some αS aggregation might be related to neuronal apoptosis or loss of nuclear membrane integrity, exposing cytosolic α-synuclein to proaggregant nuclear factors. These findings provide new clues to the pathogenesis of PD and related disorders that can lead to novel treatments of these disorders. Specifically, finding ways to limit the effects of apoptosis on αS aggregation, deposition, local uptake and subsequent propagation might significantly impact progression of disease.
Keywords: 6OHDA; Aggregation; Apoptosis; Lamin B1; MPP+; Nuclear membrane integrity; Parkinson’s disease; Proaggregant nuclear factors; α-Synuclein.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
