

Hybrid promiscuous (Hypr) GGDEF enzymes produce cyclic AMP-GMP (3', 3'-cGAMP)
- PMID: 26839412
- PMCID: PMC4763787
- DOI: 10.1073/pnas.1515287113
Hybrid promiscuous (Hypr) GGDEF enzymes produce cyclic AMP-GMP (3', 3'-cGAMP)
Abstract
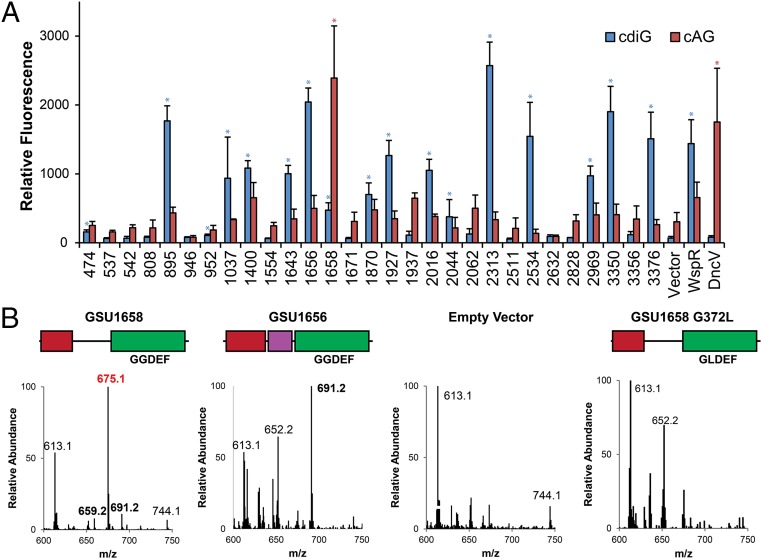
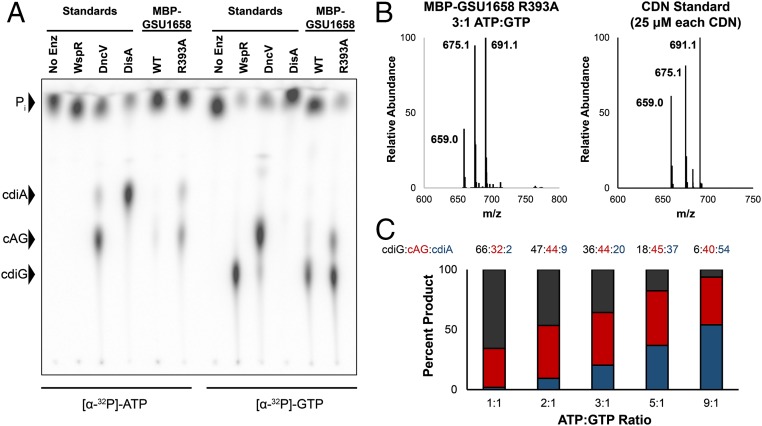
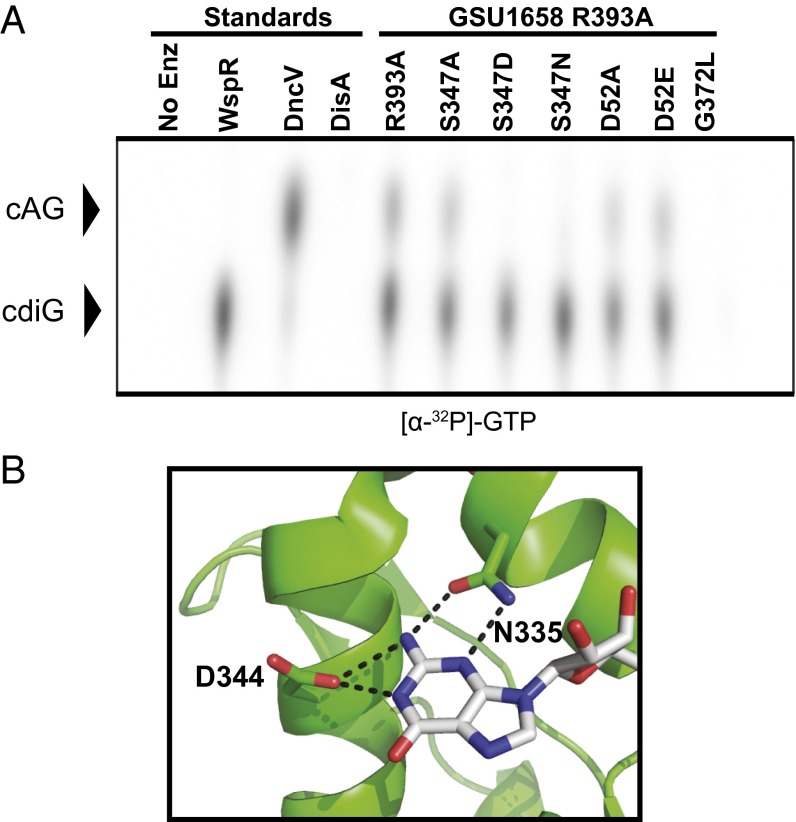
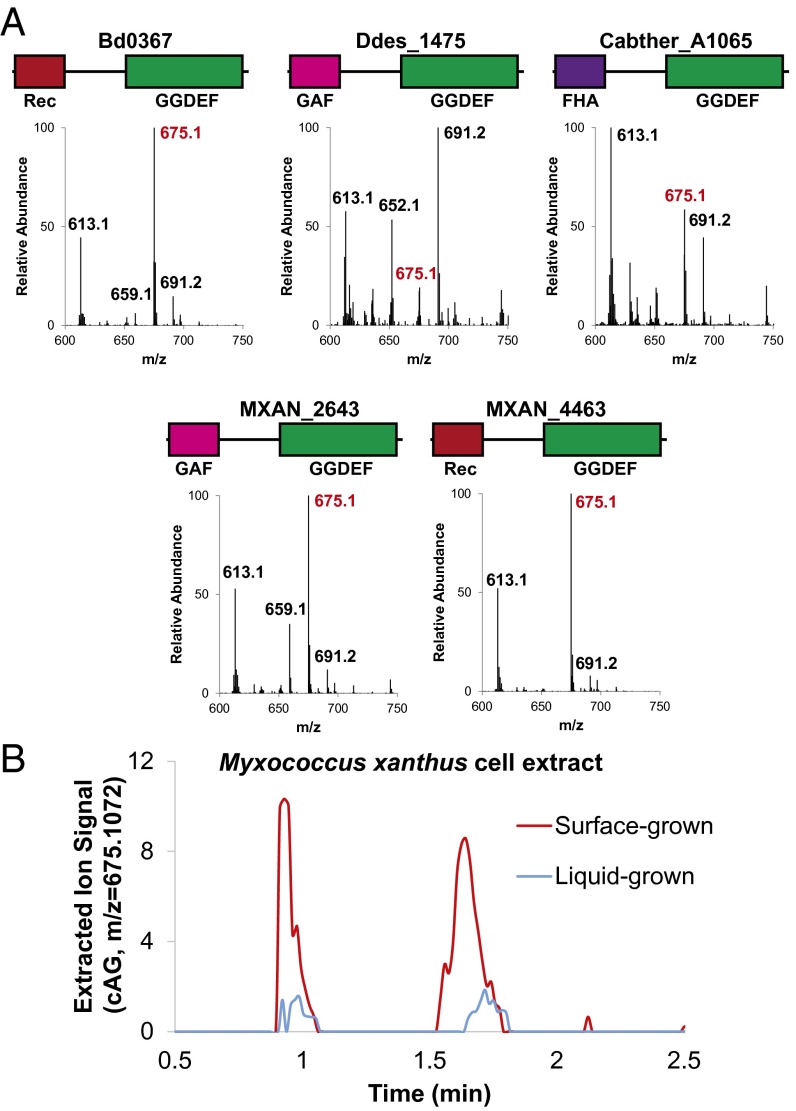
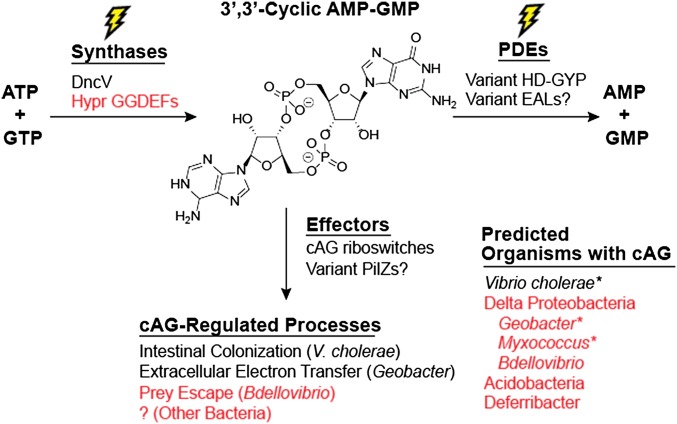
Over 30 years ago, GGDEF domain-containing enzymes were shown to be diguanylate cyclases that produce cyclic di-GMP (cdiG), a second messenger that modulates the key bacterial lifestyle transition from a motile to sessile biofilm-forming state. Since then, the ubiquity of genes encoding GGDEF proteins in bacterial genomes has established the dominance of cdiG signaling in bacteria. However, the observation that proteobacteria encode a large number of GGDEF proteins, nearing 1% of coding sequences in some cases, raises the question of why bacteria need so many GGDEF enzymes. In this study, we reveal that a subfamily of GGDEF enzymes synthesizes the asymmetric signaling molecule cyclic AMP-GMP (cAG or 3', 3'-cGAMP). This discovery is unexpected because GGDEF enzymes function as symmetric homodimers, with each monomer binding to one substrate NTP. Detailed analysis of the enzyme from Geobacter sulfurreducens showed it is a dinucleotide cyclase capable of switching the major cyclic dinucleotide (CDN) produced based on ATP-to-GTP ratios. We then establish through bioinformatics and activity assays that hybrid CDN-producing and promiscuous substrate-binding (Hypr) GGDEF enzymes are found in other deltaproteobacteria. Finally, we validated the predictive power of our analysis by showing that cAG is present in surface-grown Myxococcus xanthus. This study reveals that GGDEF enzymes make alternative cyclic dinucleotides to cdiG and expands the role of this widely distributed enzyme family to include regulation of cAG signaling.
Keywords: bacterial signaling; cyclic dinucleotides; fluorescent biosensor; second messengers; surface sensing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Structural analysis of an oxygen-regulated diguanylate cyclase.Acta Crystallogr D Biol Crystallogr. 2015 Nov;71(Pt 11):2158-77. doi: 10.1107/S139900471501545X. Epub 2015 Oct 27. Acta Crystallogr D Biol Crystallogr. 2015. PMID: 26527135
-
C-di-GMP Synthesis: Structural Aspects of Evolution, Catalysis and Regulation.J Mol Biol. 2016 Sep 25;428(19):3683-701. doi: 10.1016/j.jmb.2016.07.023. Epub 2016 Aug 4. J Mol Biol. 2016. PMID: 27498163 Review.
-
The structure and inhibition of a GGDEF diguanylate cyclase complexed with (c-di-GMP)(2) at the active site.Acta Crystallogr D Biol Crystallogr. 2011 Dec;67(Pt 12):997-1008. doi: 10.1107/S090744491104039X. Epub 2011 Nov 18. Acta Crystallogr D Biol Crystallogr. 2011. PMID: 22120736
-
Structural and Enzymatic Characterization of a cAMP-Dependent Diguanylate Cyclase from Pathogenic Leptospira Species.J Mol Biol. 2017 Jul 21;429(15):2337-2352. doi: 10.1016/j.jmb.2017.06.002. Epub 2017 Jun 7. J Mol Biol. 2017. PMID: 28601495
-
[Activity of cyclic diguanylate (c-di-GMP) in bacteria and the study of its derivatives].Yao Xue Xue Bao. 2012 Mar;47(3):307-12. Yao Xue Xue Bao. 2012. PMID: 22645753 Review. Chinese.
Cited by
-
Crystal structure and fluorescence properties of the iSpinach aptamer in complex with DFHBI.RNA. 2017 Dec;23(12):1788-1795. doi: 10.1261/rna.063008.117. Epub 2017 Sep 22. RNA. 2017. PMID: 28939697 Free PMC article.
-
Milestones in the development of Myxococcus xanthus as a model multicellular bacterium.J Bacteriol. 2025 Jul 24;207(7):e0007125. doi: 10.1128/jb.00071-25. Epub 2025 Jun 17. J Bacteriol. 2025. PMID: 40525847 Free PMC article. Review.
-
The Signaling Pathway That cGAMP Riboswitches Found: Analysis and Application of Riboswitches to Study cGAMP Signaling in Geobacter sulfurreducens.Int J Mol Sci. 2022 Jan 21;23(3):1183. doi: 10.3390/ijms23031183. Int J Mol Sci. 2022. PMID: 35163114 Free PMC article.
-
Second messengers and divergent HD-GYP phosphodiesterases regulate 3',3'-cGAMP signaling.Mol Microbiol. 2020 Jan;113(1):222-236. doi: 10.1111/mmi.14412. Epub 2019 Nov 17. Mol Microbiol. 2020. PMID: 31665539 Free PMC article.
-
Direct activation of a phospholipase by cyclic GMP-AMP in El Tor Vibrio cholerae.Proc Natl Acad Sci U S A. 2018 Jun 26;115(26):E6048-E6055. doi: 10.1073/pnas.1801233115. Epub 2018 Jun 11. Proc Natl Acad Sci U S A. 2018. PMID: 29891656 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
