

Review
doi: 10.1038/nsmb.3163.
Replication stress: getting back on track
- PMID: 26840898
- PMCID: PMC5125612
- DOI: 10.1038/nsmb.3163
Item in Clipboard
Review
Replication stress: getting back on track
Nat Struct Mol Biol.
2016 Feb.
Abstract
The replication-stress response enables the DNA replication machinery to overcome DNA lesions or intrinsic replication-fork obstacles, and it is essential to ensure faithful transmission of genetic information to daughter cells. Multiple replication stress–response pathways have been identified in recent years, thus raising questions about the specific and possibly redundant functions of these pathways. Here, we review the emerging mechanisms of the replication-stress response in mammalian cells and consider how they may influence the dynamics of the core DNA replication complex.
Figures

Different mechanisms may resume DNA synthesis when replication forks are stalled by a leading strand lesion. (a,b) Fork uncoupling/resection: Replication fork uncoupling leads to ssDNA accumulation at the fork junction through functional dissociation of the MCM helicase and the stalled polymerase. Alternatively, fork uncoupling may result from nuclease-mediated resection of stalled forks. ssDNA is rapidly coated by the single-stranded DNA binding protein RPA (yellow spheres). (c,d) Several factors, including the HDR/FA proteins BRCA1, BRCA2 and FANCD2, regulate the stability of stalled replication forks, and prevent helicase-polymerase uncoupling or nucleolytic degradation of nascent strands ,. (e) Fork repriming: DNA synthesis can be reprimed (green arrow) and reinitiated ahead of a lesion or block. The resulting gaps are repaired post-replicatively by a recombination-based mechanism or by specific Translesion Synthesis (TLS) polymerases. TLS polymerases may also function at stalled replication forks to ensure continued DNA synthesis through damaged templates (not shown). (f,g) Fork reversal: A controlled resection/uncoupling event at stalled forks promotes RAD51 loading (orange spheres) and primes fork reversal. The exact location of RAD51 binding within forks is not known. Fork reversal prevents collisions between the moving fork and a block or lesion, allowing the lesion to be repaired by the DNA repair machinery. Alternatively, it may promote lesion bypass via template switching. (h) Fork breakage: Prolonged fork stalling promotes fork cleavage by structure-specific endonucleases. Broken forks are able to resume DNA synthesis by the error-prone Break-Induced Replication (BIR) mechanism.
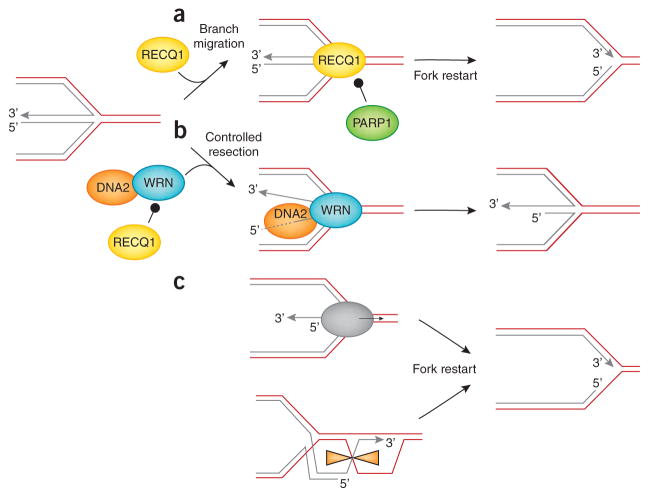
Two mechanisms of reversed replication fork resolution have been identified to date, one dependent on RECQ1 helicase and the other on DNA2 nuclease and WRN ATPase activity . (a) RECQ1 (yellow oval) restarts reversed forks via its ATPase and branch migration activity. PARP activity (green oval) is not required to form reversed forks, but it promotes the accumulation of regressed forks by inhibiting RECQ1 fork-restoration activity, thus preventing premature fork restart. (b) DNA2 and WRN (orange and green ovals, respectively) functionally interact to process reversed forks. DNA2 degrades reversed forks with a 5′ to 3′ polarity. WRN ATPase activity assists DNA2 degradation, possibly by promoting the opening of the reversed arm of the fork. RECQ1 limits DNA2 activity by an ATPase-independent mechanism. (c) Following DNA2-dependent processing, branch migration factors (grey oval) specifically recognize the partially resected reversed forks to promote fork restart. Alternatively, the newly formed 3′ overhang of the reversed fork invades the duplex ahead of the fork, resulting in a pseudo-Holliday junction structure that can be resolved by specific resolvases or dissolvases to promote fork restart.

MCM2-7 adopts an open conformation upon replication fork blockage and it is able to slide on dsDNA and bypass the ICL (indicated by a yellow star). The FANCM/MHF complex is required for this process, however the exact molecular mechanism is unclear (not shown) . The question mark indicates that it is also unclear whether CDC45 and GINS are able to bypass the roadblock in complex with MCM2-7. It is thought that the GINS proteins may be released from the complex to allow this transition . The active replisome is reestablished ahead of the block in an origin-independent fashion and the ICL is repaired post-replicatively. MCM2-7 is shown in blue, CDC45 in yellow, and GINS in red.
References
-
- Tercero JA, Diffley JF. Regulation of DNA replication fork progression through damaged DNA by the Mec1/Rad53 checkpoint. Nature. 2001;412:553–7. - PubMed
-
- De Piccoli G, et al. Replisome stability at defective DNA replication forks is independent of S phase checkpoint kinases. Mol Cell. 2012;45:696–704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
