

Somato-dendritic Synaptic Plasticity and Error-backpropagation in Active Dendrites
- PMID: 26841235
- PMCID: PMC4739747
- DOI: 10.1371/journal.pcbi.1004638
Somato-dendritic Synaptic Plasticity and Error-backpropagation in Active Dendrites
Abstract
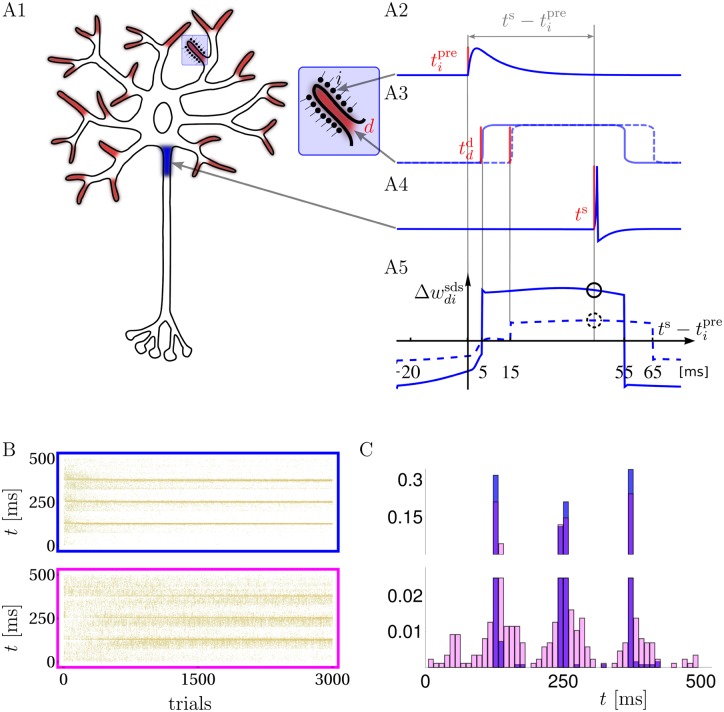
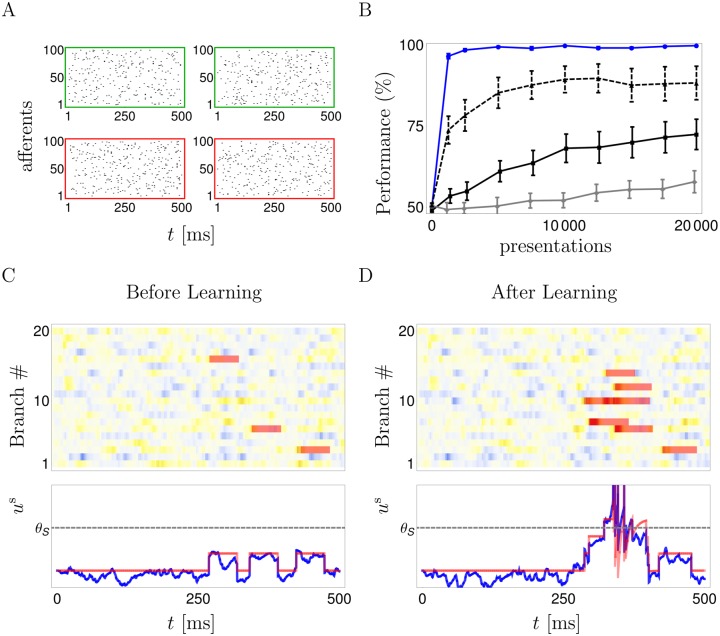
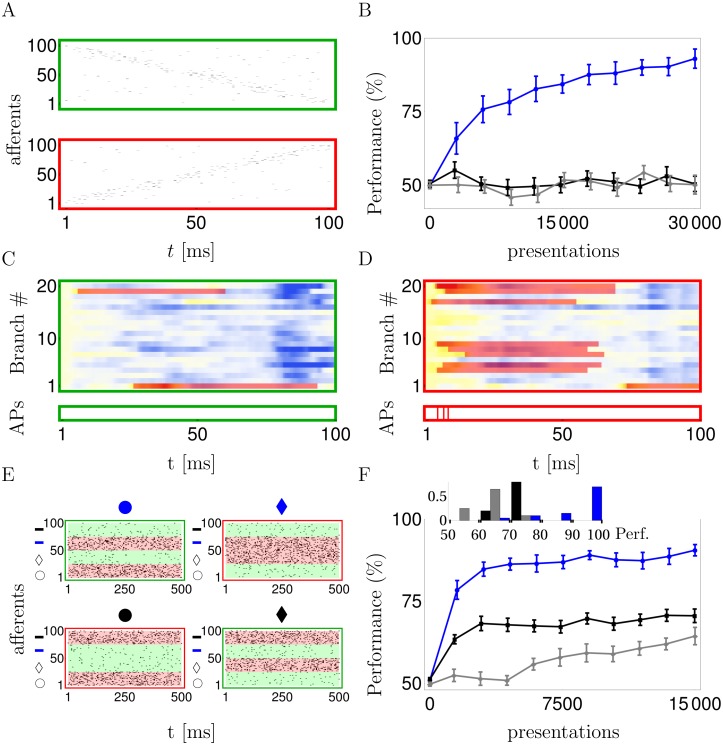
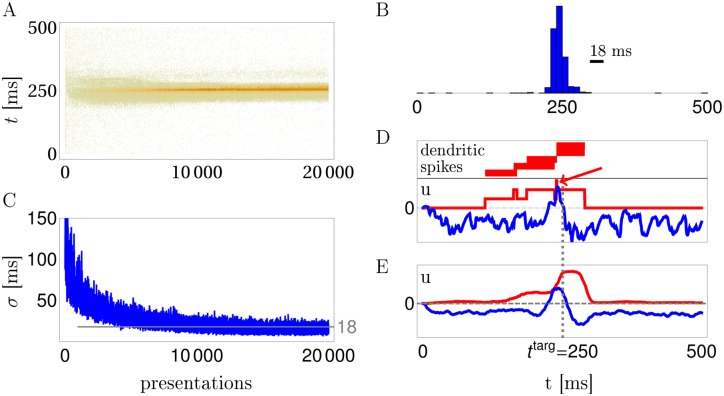
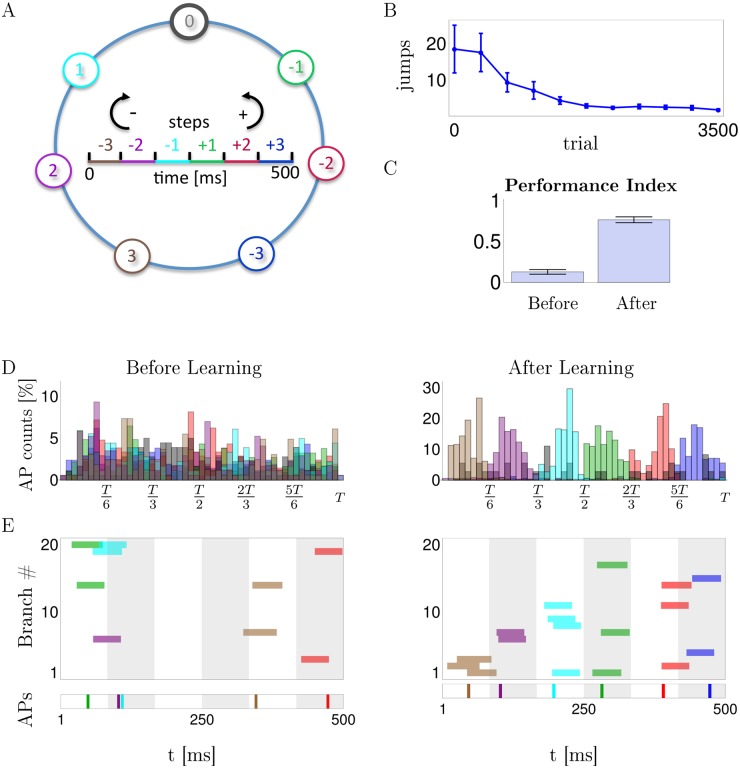
In the last decade dendrites of cortical neurons have been shown to nonlinearly combine synaptic inputs by evoking local dendritic spikes. It has been suggested that these nonlinearities raise the computational power of a single neuron, making it comparable to a 2-layer network of point neurons. But how these nonlinearities can be incorporated into the synaptic plasticity to optimally support learning remains unclear. We present a theoretically derived synaptic plasticity rule for supervised and reinforcement learning that depends on the timing of the presynaptic, the dendritic and the postsynaptic spikes. For supervised learning, the rule can be seen as a biological version of the classical error-backpropagation algorithm applied to the dendritic case. When modulated by a delayed reward signal, the same plasticity is shown to maximize the expected reward in reinforcement learning for various coding scenarios. Our framework makes specific experimental predictions and highlights the unique advantage of active dendrites for implementing powerful synaptic plasticity rules that have access to downstream information via backpropagation of action potentials.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
