

Nuclear Translocation Sequence and Region in Autographa californica Multiple Nucleopolyhedrovirus ME53 That Are Important for Optimal Baculovirus Production
- PMID: 26842471
- PMCID: PMC4810555
- DOI: 10.1128/JVI.03115-15
Nuclear Translocation Sequence and Region in Autographa californica Multiple Nucleopolyhedrovirus ME53 That Are Important for Optimal Baculovirus Production
Abstract
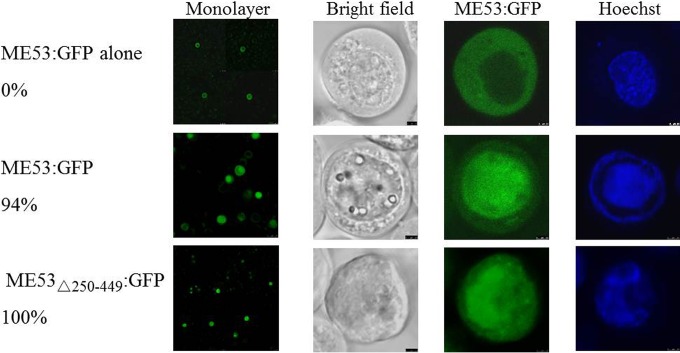
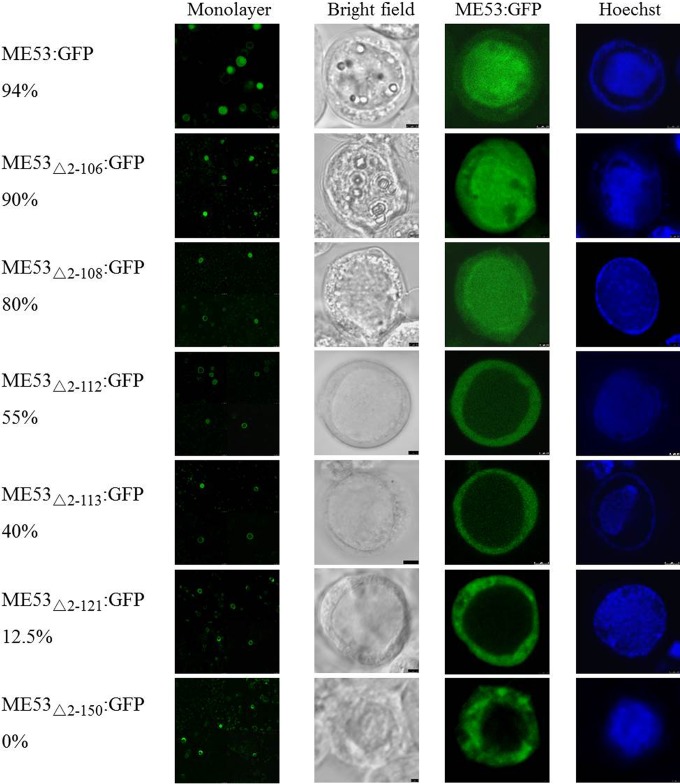
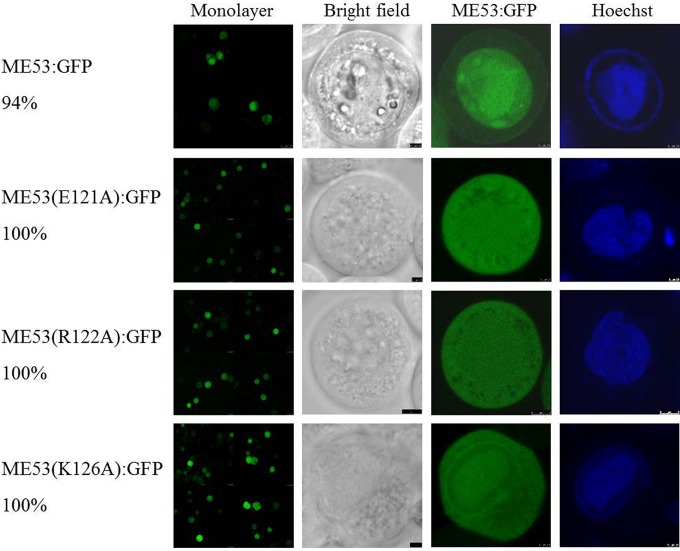
Autographa californicamultiple nucleopolyhedrovirus (AcMNPV) is in the familyBaculoviridae, genusAlphabaculovirus AcMNPVme53is a highly conserved immediate early gene in all lepidopteran baculoviruses that have been sequenced and is transcribed up to late times postinfection. Althoughme53is not essential for viral DNA synthesis, infectious budded virus (BV) production is greatly attenuated when it is deleted. ME53 associates with the nucleocapsid on both budded virus and occlusion-derived virus, but not with the virus envelope. ME53 colocalizes in plasma membrane foci with the envelope glycoprotein GP64 in a GP64-dependent manner. ME53 localizes in the cytoplasm early postinfection, and despite the lack of a reported nuclear localization signal (NLS), ME53 translocates to the nucleus at late times postinfection. To map determinants of ME53 that facilitate its nuclear translocation, recombinant AcMNPV bacmids containing a series of ME53 truncations, internal deletions, and peptides fused with hemagglutinin (HA) or green fluorescent protein (GFP) tags were constructed. Intracellular-localization studies identified residues within amino acids 109 to 137 at the N terminus of ME53 that acted as the nuclear translocation sequence (NTS), facilitating its nuclear transport at late times postinfection. The first 100 N-terminal amino acids and the last 50 C-terminal amino acids of ME53 are dispensable for high levels of budded virus production. The region within amino acids 101 to 398, which also contains the NTS, is critical for optimal levels of budded virus production.
Importance: Baculovirusme53is a conserved immediate early gene found in all sequenced lepidopteran alpha- and betabaculoviruses. We first identified residues within amino acids 109 to 137 at the N terminus that act as the ME53 nuclear translocation sequence (NTS) to facilitate its nuclear translocation and defined an internal region within amino acids 101 to 398, which includes the NTS, as being necessary for optimal budded virus production. Altogether, these results indicate a previously unidentified nuclear role that ME53 plays in virus replication.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures







Similar articles
-
me53 encoded by Autographa californica multiple nucleopolyhedrovirus: from mechanism to function.Virus Genes. 2023 Apr;59(2):188-194. doi: 10.1007/s11262-022-01943-3. Epub 2022 Oct 13. Virus Genes. 2023. PMID: 36229721 Review.
-
Autographa californica multiple nucleopolyhedrovirus me53 (ac140) is a nonessential gene required for efficient budded-virus production.J Virol. 2009 Aug;83(15):7440-8. doi: 10.1128/JVI.02390-08. Epub 2009 May 20. J Virol. 2009. PMID: 19457997 Free PMC article.
-
Immediate-early protein ME53 forms foci and colocalizes with GP64 and the major capsid protein VP39 at the cell membranes of Autographa californica multiple nucleopolyhedrovirus-infected cells.J Virol. 2011 Oct;85(19):9696-707. doi: 10.1128/JVI.00833-11. Epub 2011 Jul 20. J Virol. 2011. PMID: 21775466 Free PMC article.
-
Characterization of AcMNPV with a deletion of me53 gene.Virus Genes. 2007 Apr;34(2):223-32. doi: 10.1007/s11262-006-0045-0. Epub 2006 Nov 10. Virus Genes. 2007. PMID: 17096186
-
Autographa californica multiple nucleopolyhedrovirus DNA polymerase C terminus is required for nuclear localization and viral DNA replication.J Virol. 2014 Sep;88(18):10918-33. doi: 10.1128/JVI.01167-14. Epub 2014 Jul 9. J Virol. 2014. PMID: 25008932 Free PMC article.
Cited by
-
A CRM1-dependent nuclear export signal in Autographa californica multiple nucleopolyhedrovirus Ac93 is important for the formation of intranuclear microvesicles.J Virol. 2024 May 14;98(5):e0029924. doi: 10.1128/jvi.00299-24. Epub 2024 Apr 1. J Virol. 2024. PMID: 38557225 Free PMC article.
-
Experimental and evolutionary evidence for horizontal transfer of an envelope fusion protein gene between thogotoviruses and baculoviruses.J Virol. 2025 Jul 22;99(7):e0214824. doi: 10.1128/jvi.02148-24. Epub 2025 Jun 25. J Virol. 2025. PMID: 40558095 Free PMC article.
-
Mechanisms Mediating Nuclear Trafficking Involved in Viral Propagation by DNA Viruses.Viruses. 2019 Nov 7;11(11):1035. doi: 10.3390/v11111035. Viruses. 2019. PMID: 31703327 Free PMC article. Review.
-
Protein composition analysis of polyhedra matrix of Bombyx mori nucleopolyhedrovirus (BmNPV) showed powerful capacity of polyhedra to encapsulate foreign proteins.Sci Rep. 2017 Aug 18;7(1):8768. doi: 10.1038/s41598-017-08987-8. Sci Rep. 2017. PMID: 28821766 Free PMC article.
-
me53 encoded by Autographa californica multiple nucleopolyhedrovirus: from mechanism to function.Virus Genes. 2023 Apr;59(2):188-194. doi: 10.1007/s11262-022-01943-3. Epub 2022 Oct 13. Virus Genes. 2023. PMID: 36229721 Review.
References
-
- Funk J, Braunagel SC, Rohrmann GF. 1997. Baculovirus structure, p 7–32. Miller LK, () The baculoviruses. Plenum Press, New York, NY.
-
- Federici BA. 1997. Baculovirus pathogenesis, p 33–60. In Miller LK. (ed), The baculoviruses. Plenum Press, New York, NY.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
