

Cancer-associated SF3B1 mutations affect alternative splicing by promoting alternative branchpoint usage
- PMID: 26842708
- PMCID: PMC4743009
- DOI: 10.1038/ncomms10615
Cancer-associated SF3B1 mutations affect alternative splicing by promoting alternative branchpoint usage
Abstract
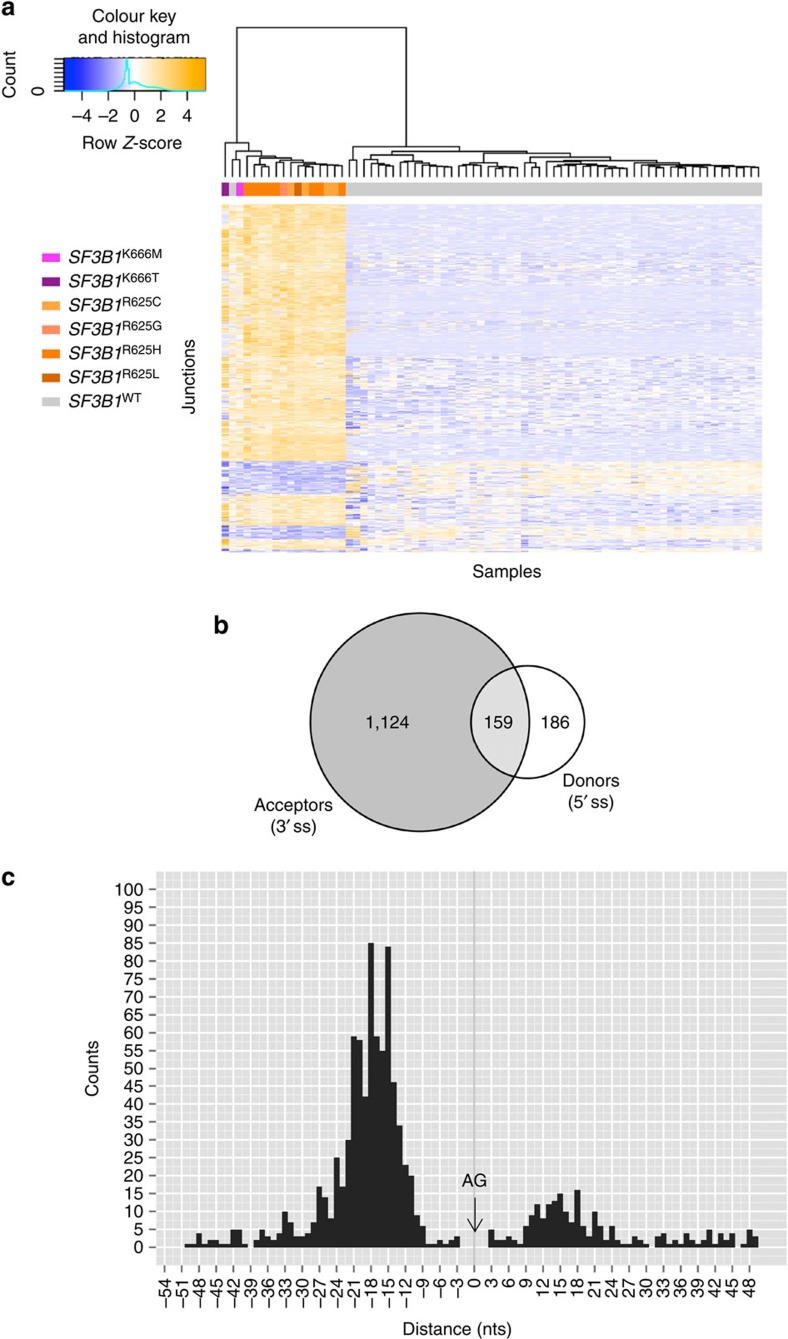
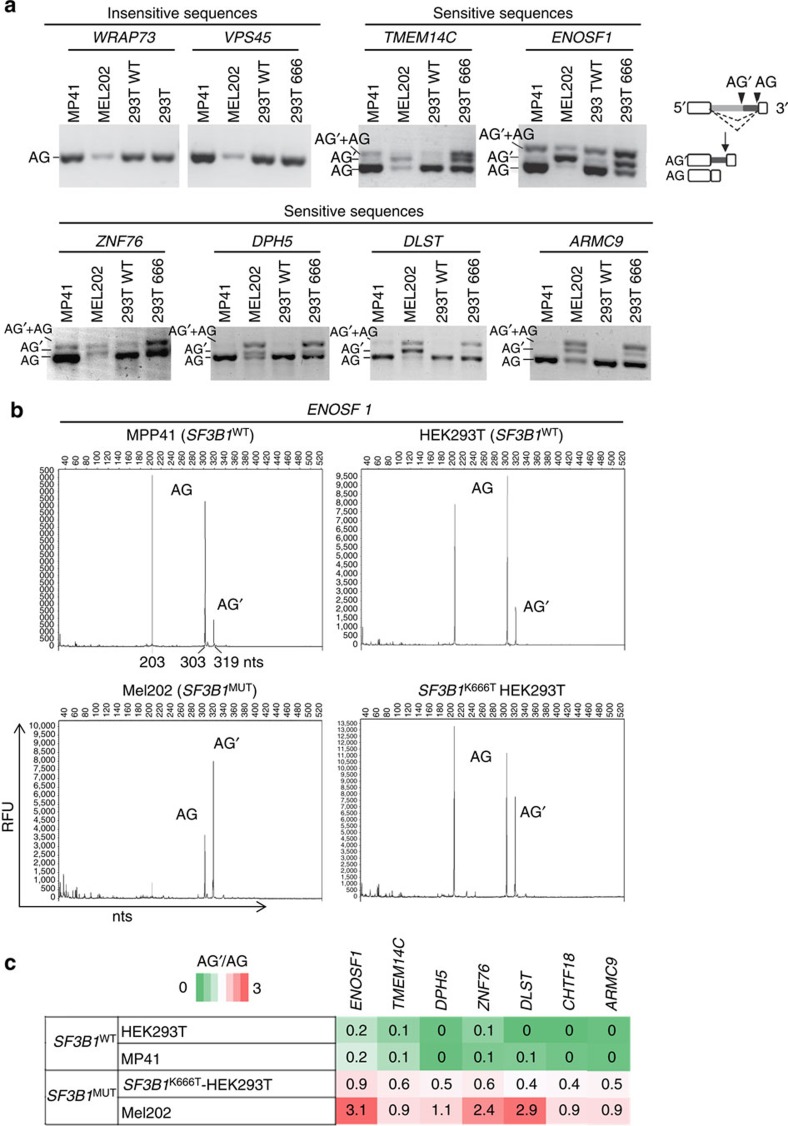
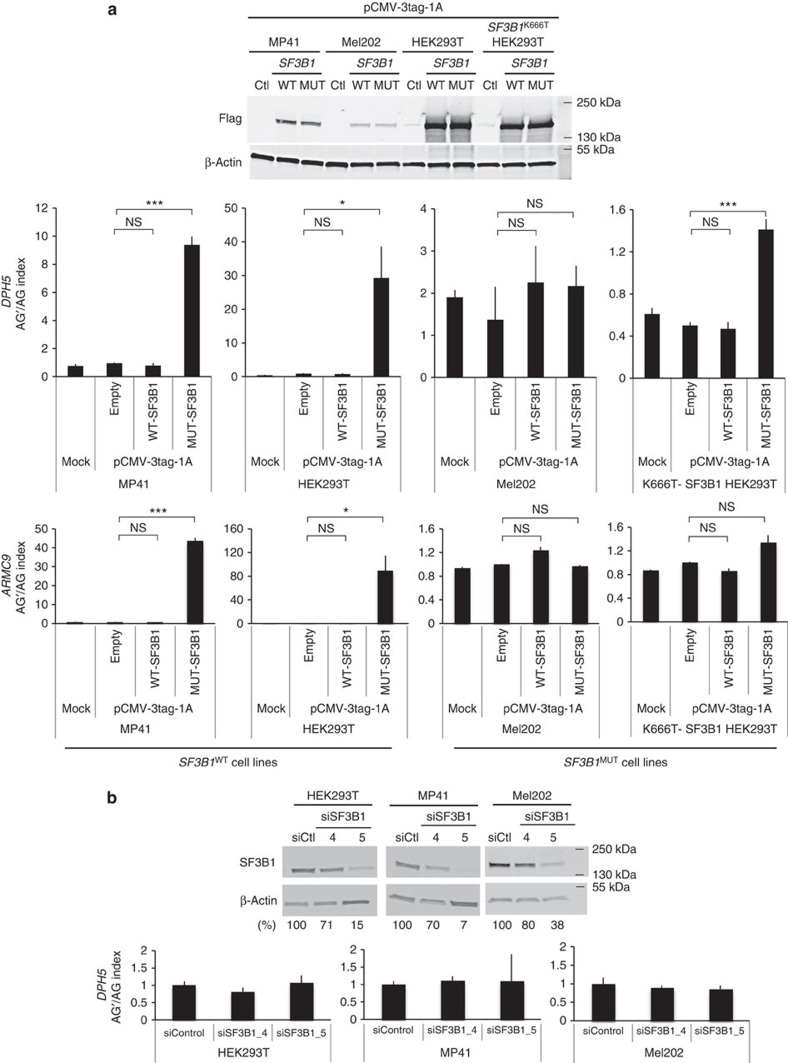
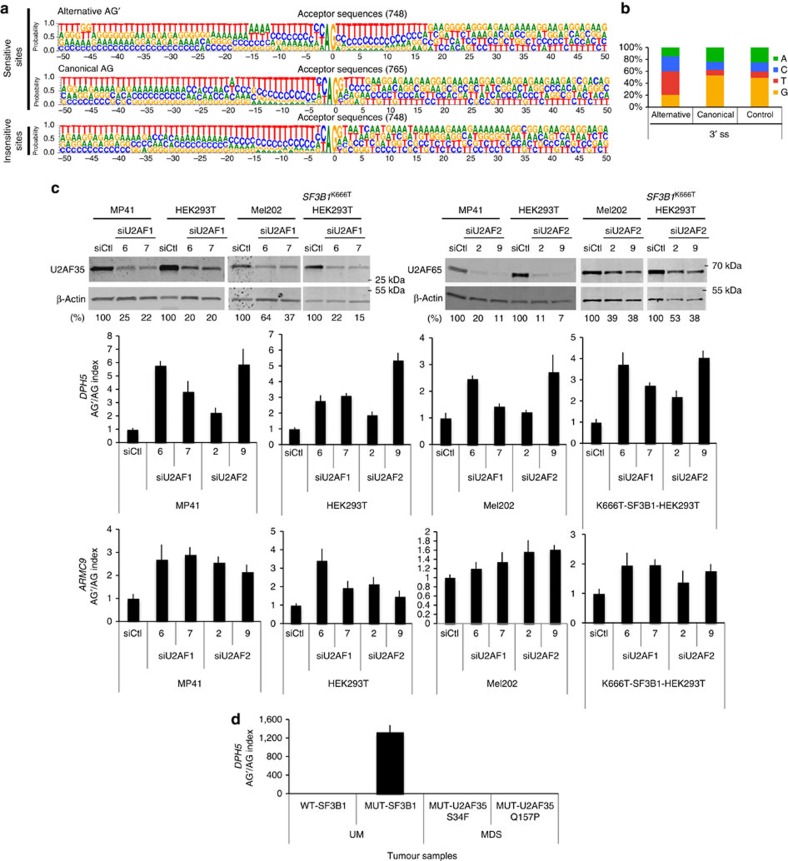
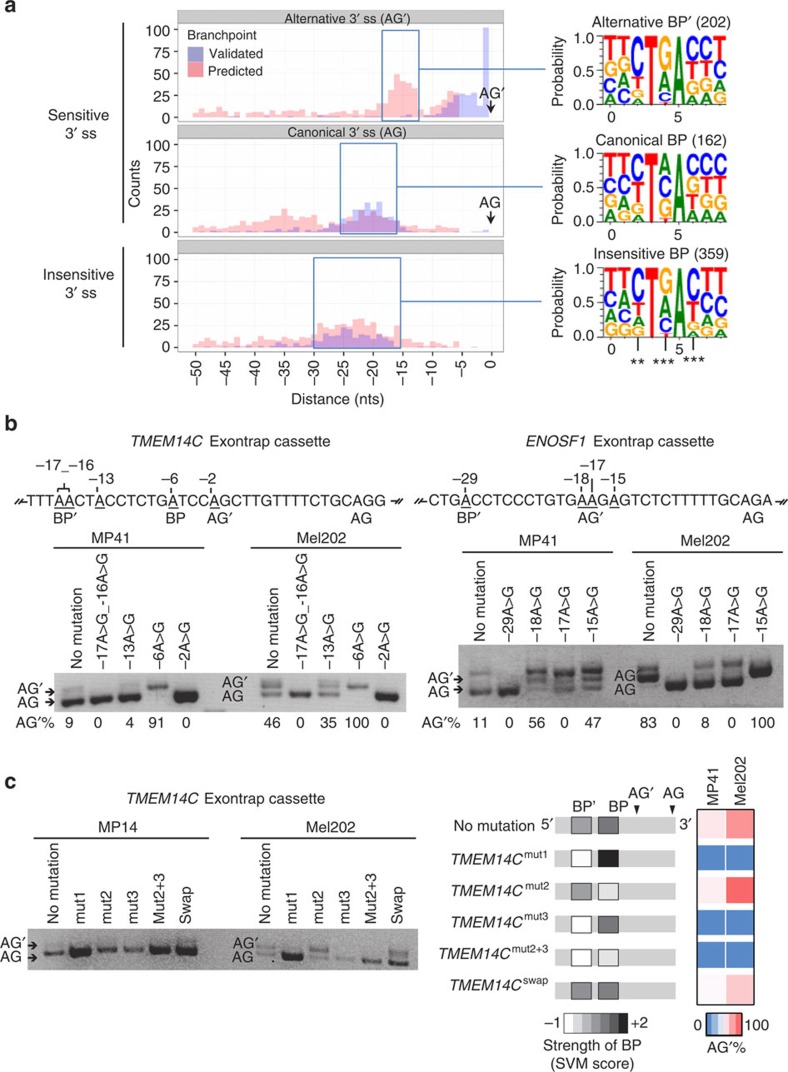
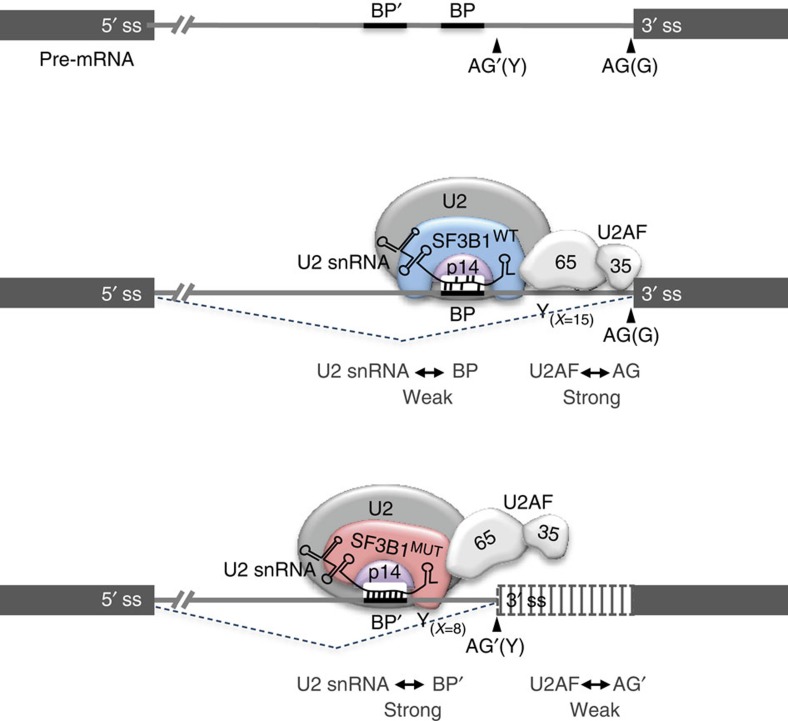
Hotspot mutations in the spliceosome gene SF3B1 are reported in ∼20% of uveal melanomas. SF3B1 is involved in 3'-splice site (3'ss) recognition during RNA splicing; however, the molecular mechanisms of its mutation have remained unclear. Here we show, using RNA-Seq analyses of uveal melanoma, that the SF3B1(R625/K666) mutation results in deregulated splicing at a subset of junctions, mostly by the use of alternative 3'ss. Modelling the differential junctions in SF3B1(WT) and SF3B1(R625/K666) cell lines demonstrates that the deregulated splice pattern strictly depends on SF3B1 status and on the 3'ss-sequence context. SF3B1(WT) knockdown or overexpression do not reproduce the SF3B1(R625/K666) splice pattern, qualifying SF3B1(R625/K666) as change-of-function mutants. Mutagenesis of predicted branchpoints reveals that the SF3B1(R625/K666)-promoted splice pattern is a direct result of alternative branchpoint usage. Altogether, this study provides a better understanding of the mechanisms underlying splicing alterations induced by mutant SF3B1 in cancer, and reveals a role for alternative branchpoints in disease.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
