

Zebrafish as a Model to Investigate Dynamin 2-Related Diseases
- PMID: 26842864
- PMCID: PMC4740890
- DOI: 10.1038/srep20466
Zebrafish as a Model to Investigate Dynamin 2-Related Diseases
Abstract
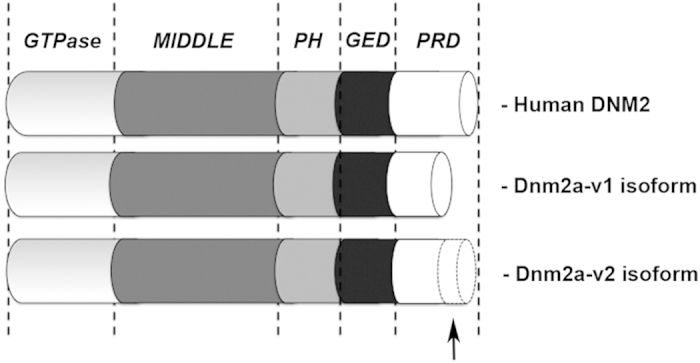
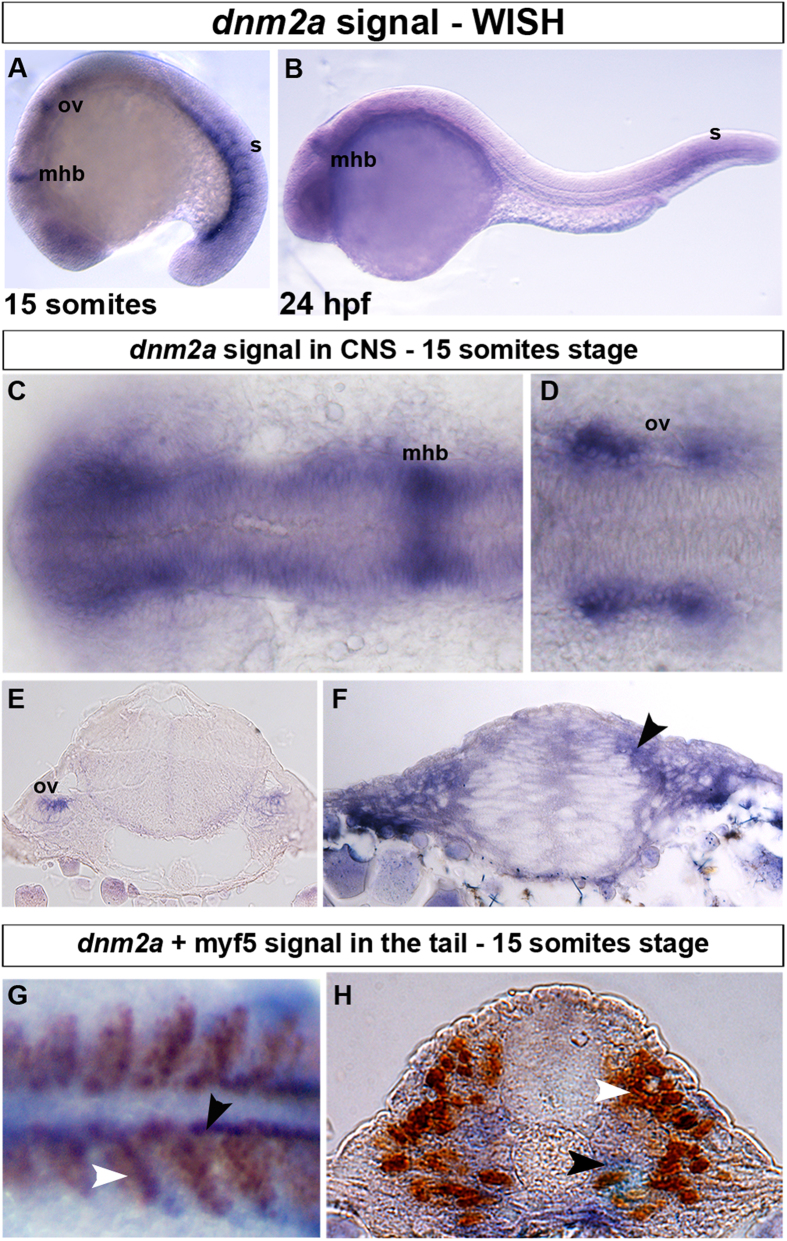
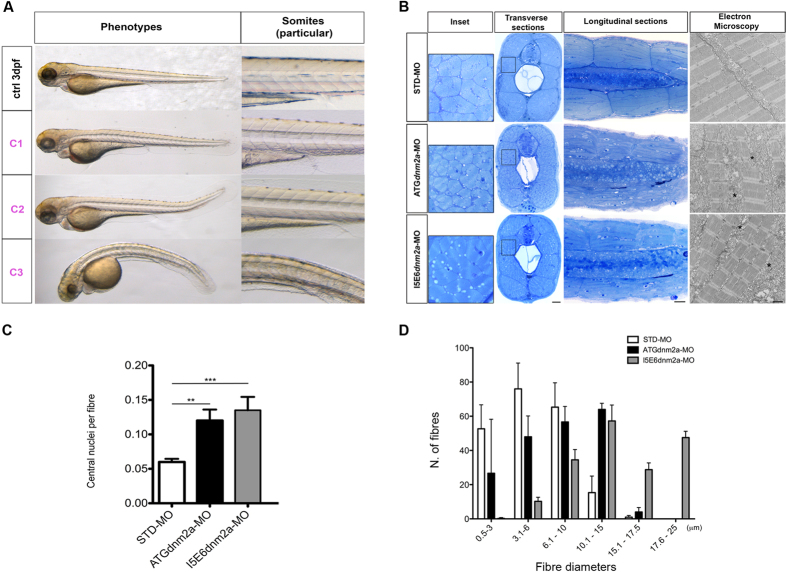
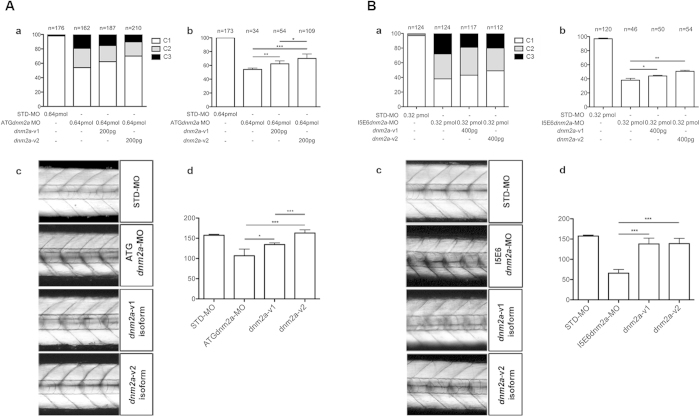
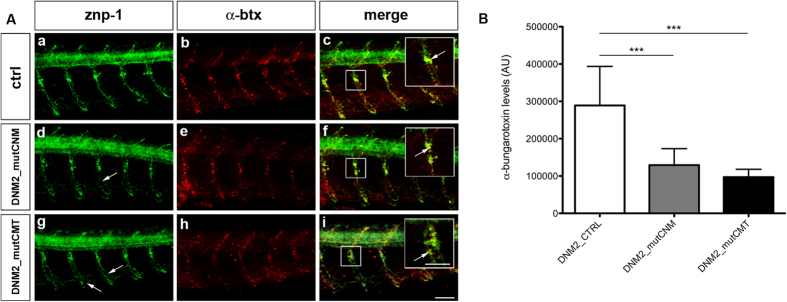
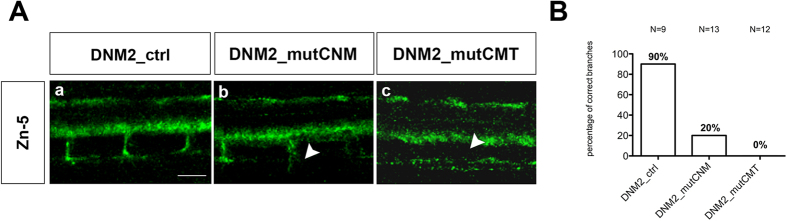
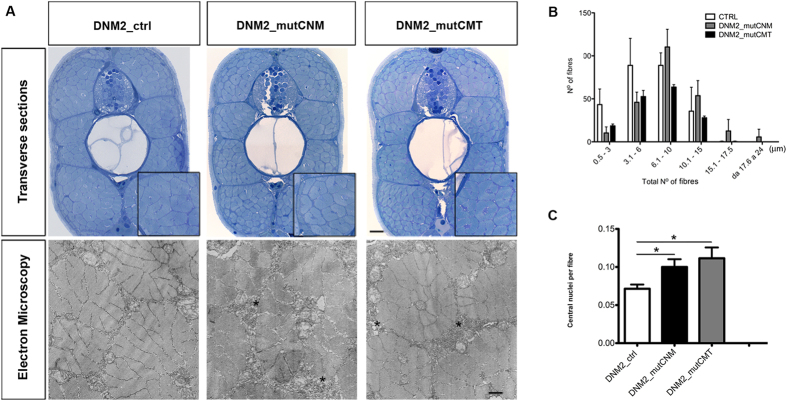
Mutations in the dynamin-2 gene (DNM2) cause autosomal dominant centronuclear myopathy (CNM) and dominant intermediate Charcot-Marie-Tooth (CMT) neuropathy type B (CMTDIB). As the relation between these DNM2-related diseases is poorly understood, we used zebrafish to investigate the effects of two different DNM2 mutations. First we identified a new alternatively spliced zebrafish dynamin-2a mRNA (dnm2a-v2) with greater similarity to human DNM2 than the deposited sequence. Then we knocked-down the zebrafish dnm2a, producing defects in muscle morphology. Finally, we expressed two mutated DNM2 mRNA by injecting zebrafish embryos with human mRNAs carrying the R522H mutation, causing CNM, or the G537C mutation, causing CMT. Defects arose especially in secondary motor neuron formation, with incorrect branching in embryos injected with CNM-mutated mRNA, and total absence of branching in those injected with CMT-mutated mRNA. Muscle morphology in embryos injected with CMT-mutated mRNA appeared less regularly organized than in those injected with CNM-mutated mRNA. Our results showing, a continuum between CNM and CMTDIB phenotypes in zebrafish, similarly to the human conditions, confirm this animal model to be a powerful tool to investigate mutations of DNM2 in vivo.
Figures
References
-
- Dyck P. J., Lebo R. & Carney J. A. Hereditary motor and sensory neuropathies. Peripheral neuropathy 3rd edn (eds Poduslo J. F. et al. ) 1094–1136 (Saunders, 1993).
-
- Nagamatsu M., Jenkins R. B., Shaid D. J., Klein D. M. & Dyck P. J. Hereditary Motor and Sensory Neuropathy Type 2C is genetically distinct from types 2B and 2D. Arch. Neurol. 57, 669–672 (2000). - PubMed
-
- Berger P., Young P. & Suter U. Molecular cell Biology of Charcot-Marie-Tooth disease. Neurogen. 4, 1–15 (2002). - PubMed
-
- Züchner S. et al. Mutations in the pleckstrin homology domain of dynamin 2 cause dominant intermediate Charcot-Marie-Tooth disease. Nat. Genet. 37, 289–294 (2005). - PubMed
-
- Praefcke G. J. K. & McMahon H. T. The dynamin superfamily: universal membrane tubulation and fission molecules? Nat. Rev. | mol. cell. bio. 5, 133–147 (2004). - PubMed
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
