

Engineered Tug-of-War Between Kinesin and Dynein Controls Direction of Microtubule Based Transport In Vivo
- PMID: 26843027
- PMCID: PMC5578396
- DOI: 10.1111/tra.12385
Engineered Tug-of-War Between Kinesin and Dynein Controls Direction of Microtubule Based Transport In Vivo
Abstract
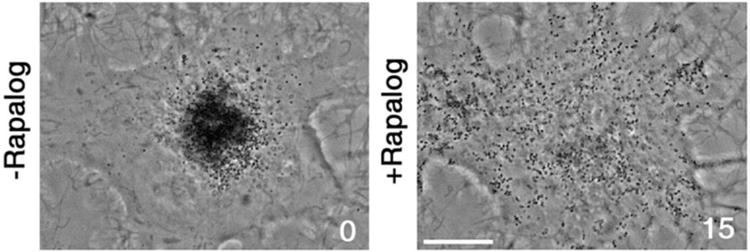
Bidirectional transport of membrane organelles along microtubules (MTs) is driven by plus-end directed kinesins and minus-end directed dynein bound to the same cargo. Activities of opposing MT motors produce bidirectional movement of membrane organelles and cytoplasmic particles along MT transport tracks. Directionality of MT-based transport might be controlled by a protein complex that determines which motor type is active at any given moment of time, or determined by the outcome of a tug-of-war between MT motors dragging cargo organelles in opposite directions. However, evidence in support of each mechanisms of regulation is based mostly on the results of theoretical analyses or indirect experimental data. Here, we test whether the direction of movement of membrane organelles in vivo can be controlled by the tug-of-war between opposing MT motors alone, by attaching a large number of kinesin-1 motors to organelles transported by dynein to minus-ends of MTs. We find that recruitment of kinesin significantly reduces the length and velocity of minus-end-directed dynein-dependent MT runs, leading to a reversal of the overall direction of dynein-driven organelles in vivo. Therefore, in the absence of external regulators tug-of-war between opposing MT motors alone is sufficient to determine the directionality of MT transport in vivo.
Keywords: dynein; intracellular transport; kinesin; melanophore; pigment granule.
© 2016 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Figures
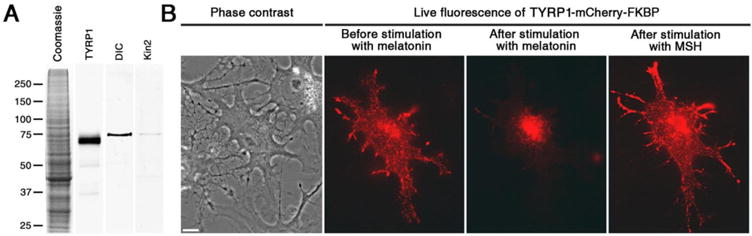
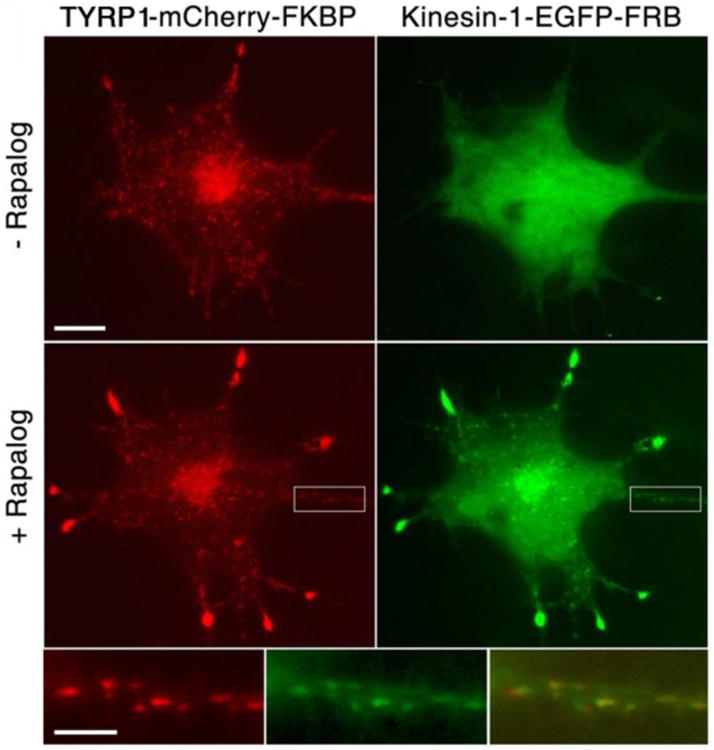
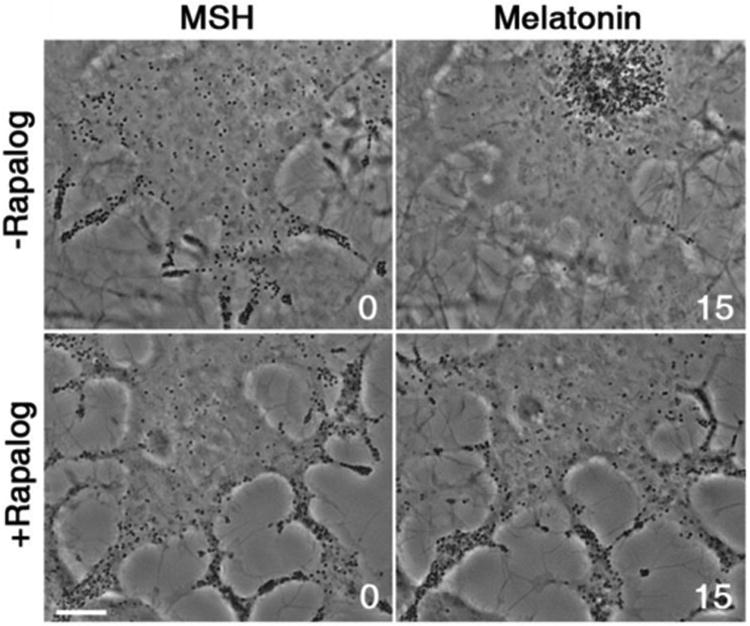
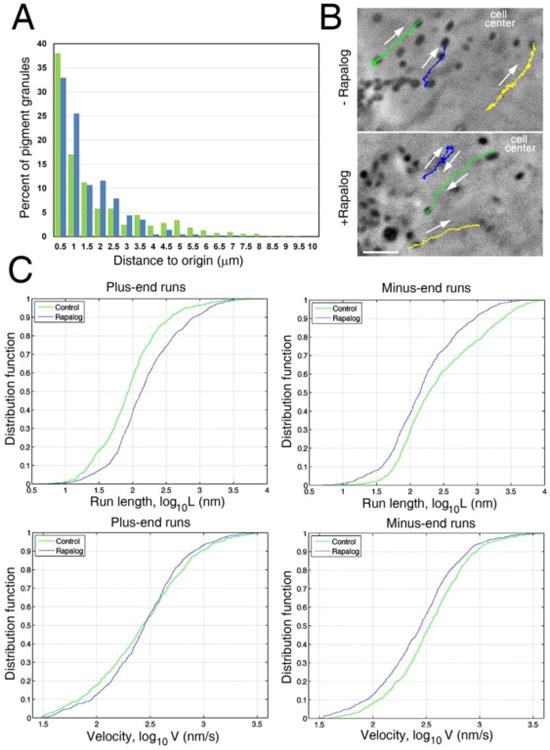
References
-
- Vale RD. The molecular motor toolbox for intracellular transport. Cell. 2003;112(4):467–480. - PubMed
-
- Gross SP. Hither and yon: a review of bi-directional microtubule-based transport. Phys Biol. 2004;1(1-2):R1–11. - PubMed
-
- Welte MA. Bidirectional transport along microtubules. Curr Biol. 2004;14(13):R525–537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
