

Orexin Neurons Respond Differentially to Auditory Cues Associated with Appetitive versus Aversive Outcomes
- PMID: 26843654
- PMCID: PMC4737782
- DOI: 10.1523/JNEUROSCI.3903-15.2016
Orexin Neurons Respond Differentially to Auditory Cues Associated with Appetitive versus Aversive Outcomes
Abstract
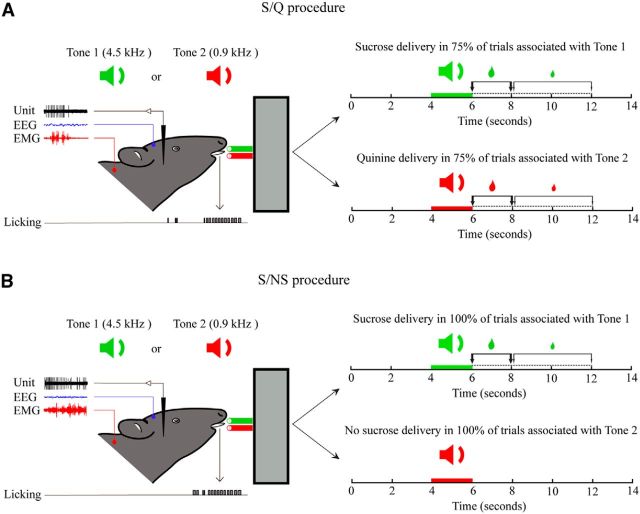
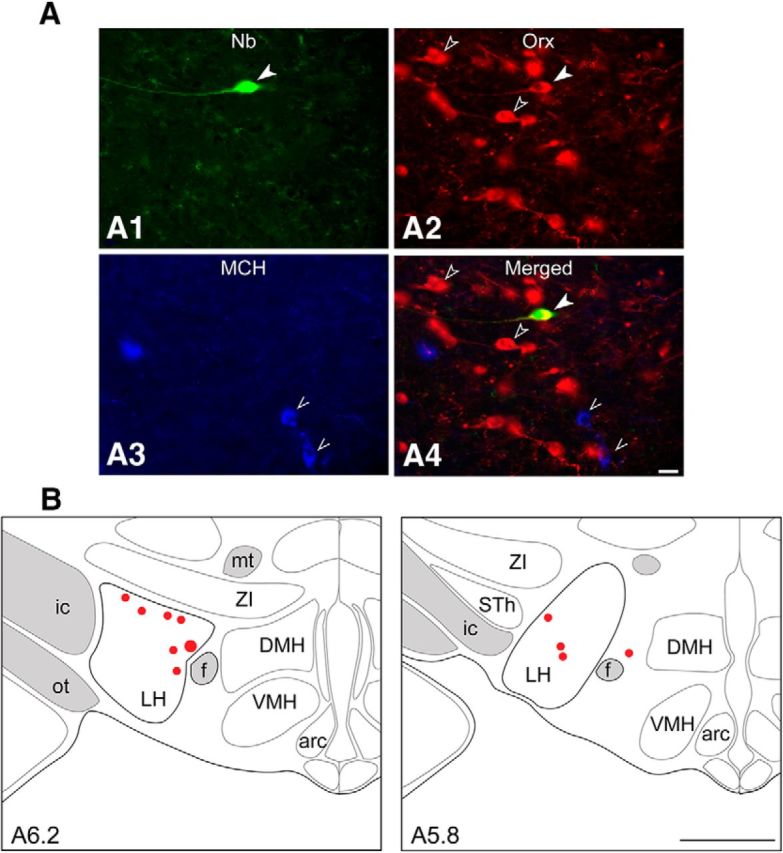
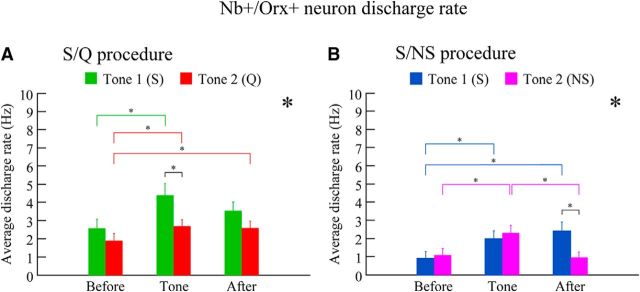
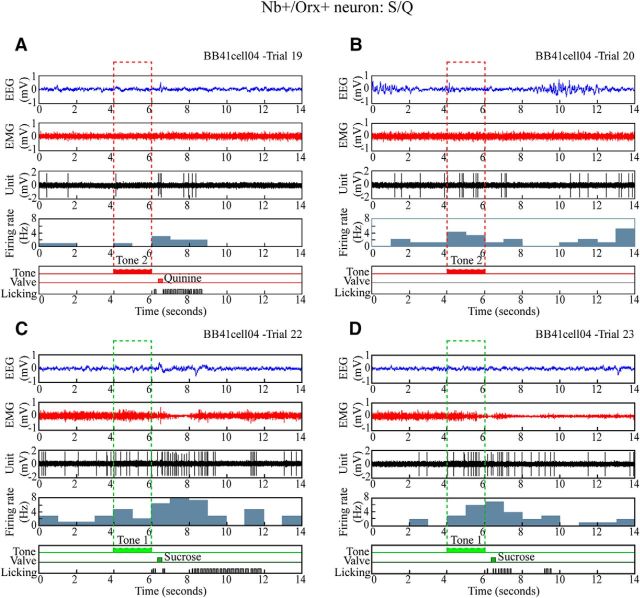
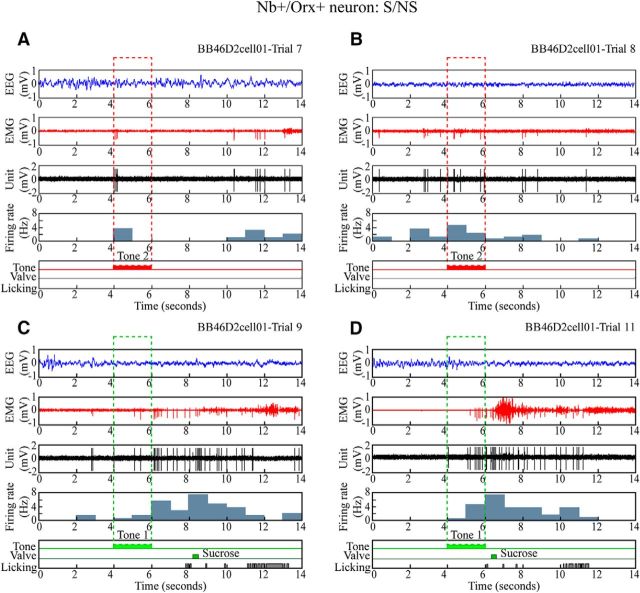
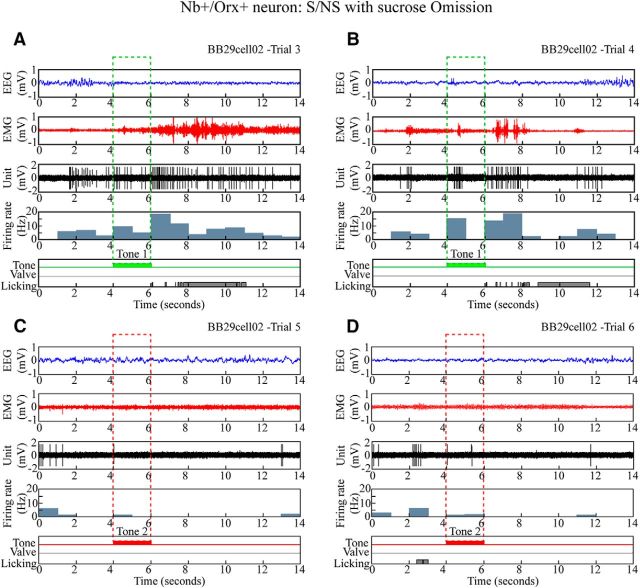
Orexin (Orx) neurons are known to be involved in the promotion and maintenance of waking because they discharge in association with cortical activation and muscle tone during waking and because, in their absence, waking with muscle tone cannot be maintained and narcolepsy with cataplexy ensues. Whether Orx neurons discharge during waking in association with particular conditions, notably with appetitive versus aversive stimuli or positive versus negative emotions, is debated and considered important in understanding their role in supporting particular waking behaviors. Here, we used the technique of juxtacellular recording and labeling in head-fixed rats to characterize the discharge of Orx neurons during the performance of an associative discrimination task with auditory cues for appetitive versus aversive outcomes. Of 57 active, recorded, and neurobiotin-labeled neurons in the lateral hypothalamus, 11 were immunohistochemically identified as Orx-positive (Orx(+)), whereas none were identified as melanin-concentrating hormone-positive. Orx(+) neurons discharged at significantly higher rates during the tone associated with sucrose than during the tone associated with quinine delivered upon licking. They also discharged at high rates after the tone associated with sucrose. Across periods and outcomes, their discharge was positively correlated with EEG gamma activity and EMG activity, which is indicative of cortical activation and behavioral arousal. These results suggest that Orx neurons discharge in a manner characteristic of reward neurons yet also characteristic of arousal neurons. Accordingly, the Orx neurons may respond to and participate in reward processes while modulating cortical activity and muscle tone to promote and maintain arousal along with learned adaptive behavioral responses.
Significance statement: Orexin neurons play a critical role in promoting and maintaining a waking state because, in their absence, narcolepsy with cataplexy ensues. Known to discharge during waking and not during sleep, they have also been proposed to be selectively active during appetitive behaviors. Here, we recorded and labeled neurons in rats to determine the discharge of immunohistochemically identified orexin neurons during performance of an associative discrimination task. Orexin neurons responded differentially to auditory cues associated with appetitive sucrose versus aversive quinine, indicating that they behave like reward neurons. However, correlated discharge with cortical and muscle activity indicates that they also behave like arousal neurons and can thus promote cortical activation with behavioral arousal and muscle tone during adaptive waking behaviors.
Keywords: EEG; EMG; MCH; arousal; reward; sleep.
Copyright © 2016 the authors 0270-6474/16/361747-11$15.00/0.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources