

Pet-1 Switches Transcriptional Targets Postnatally to Regulate Maturation of Serotonin Neuron Excitability
- PMID: 26843655
- PMCID: PMC4737783
- DOI: 10.1523/JNEUROSCI.3798-15.2016
Pet-1 Switches Transcriptional Targets Postnatally to Regulate Maturation of Serotonin Neuron Excitability
Abstract
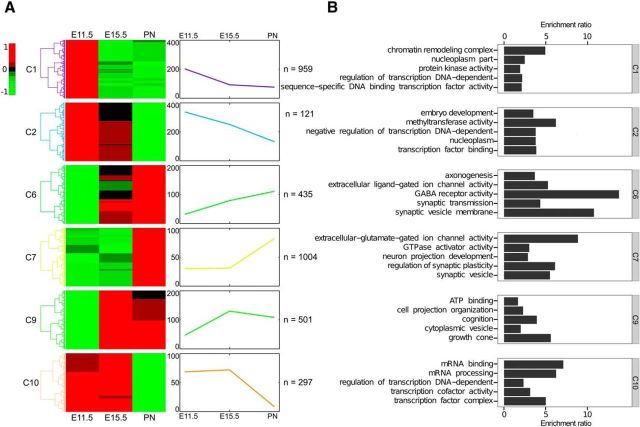
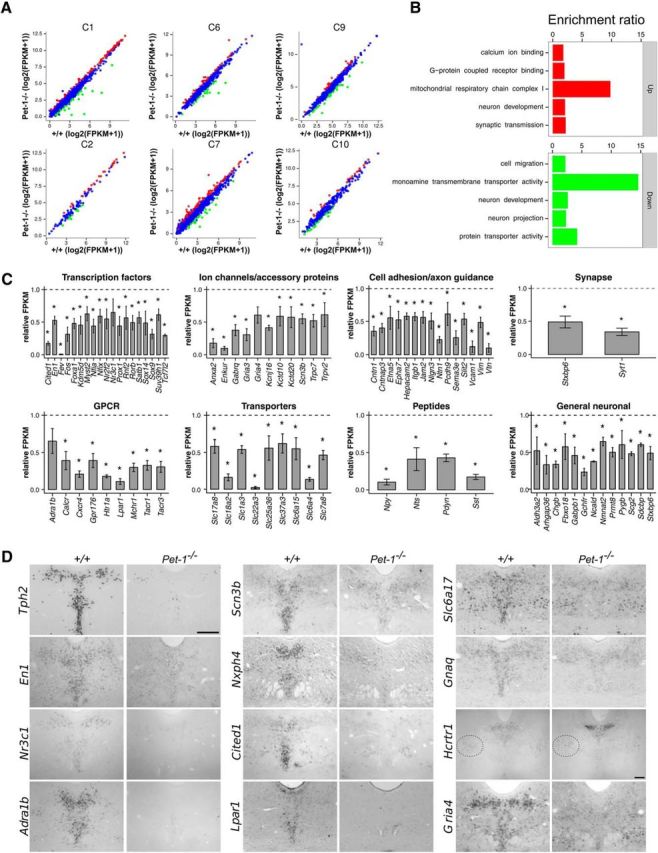
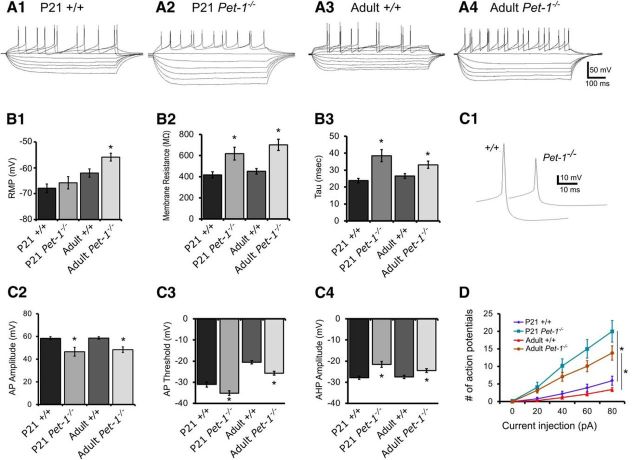
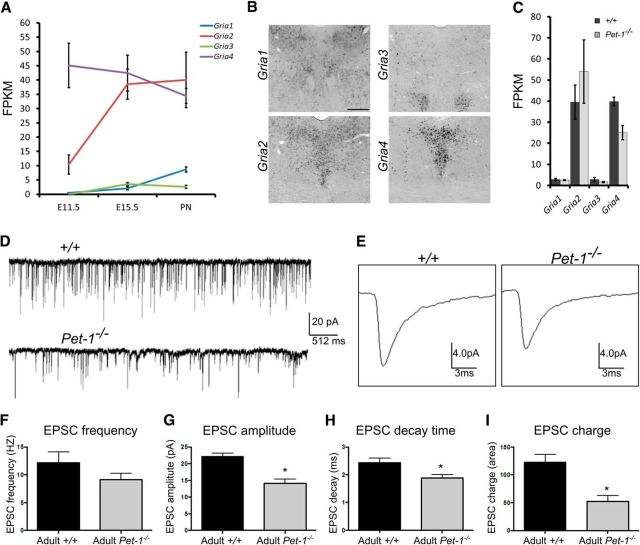
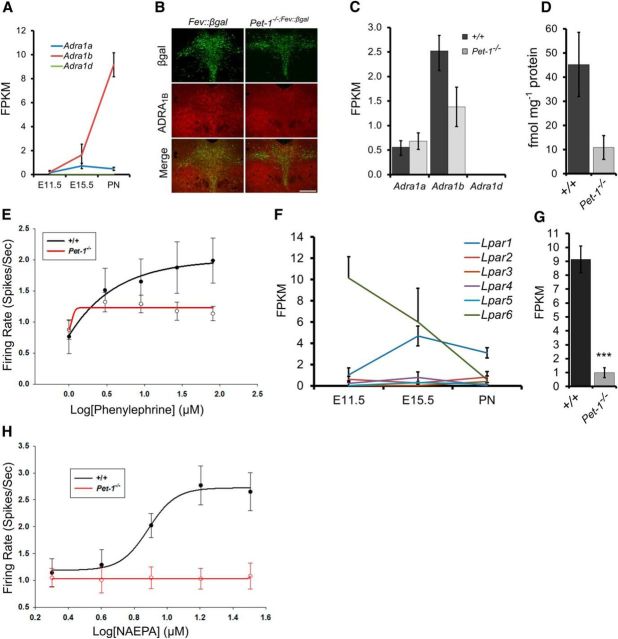
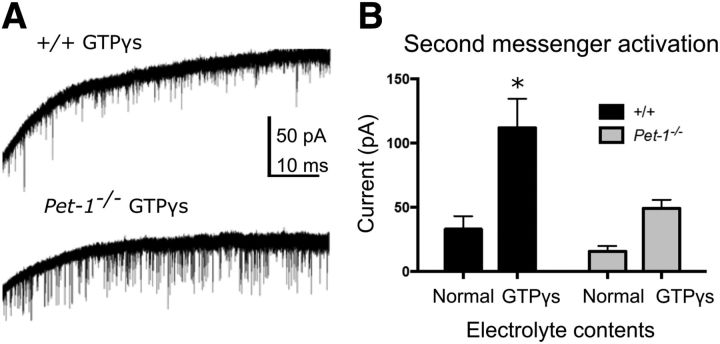
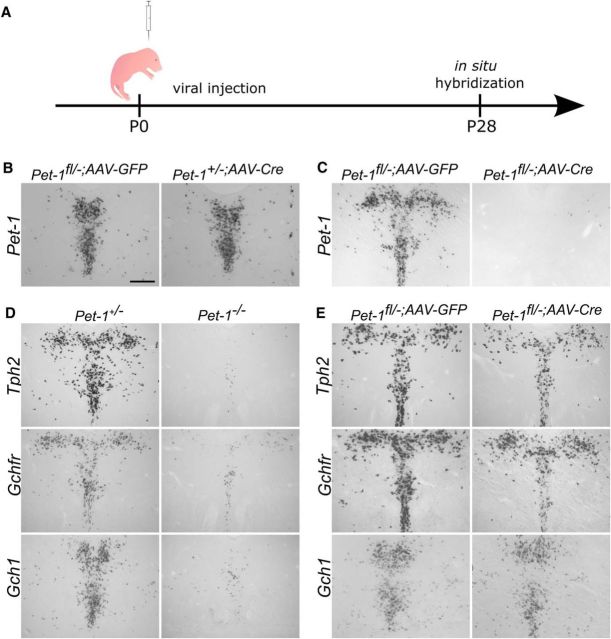
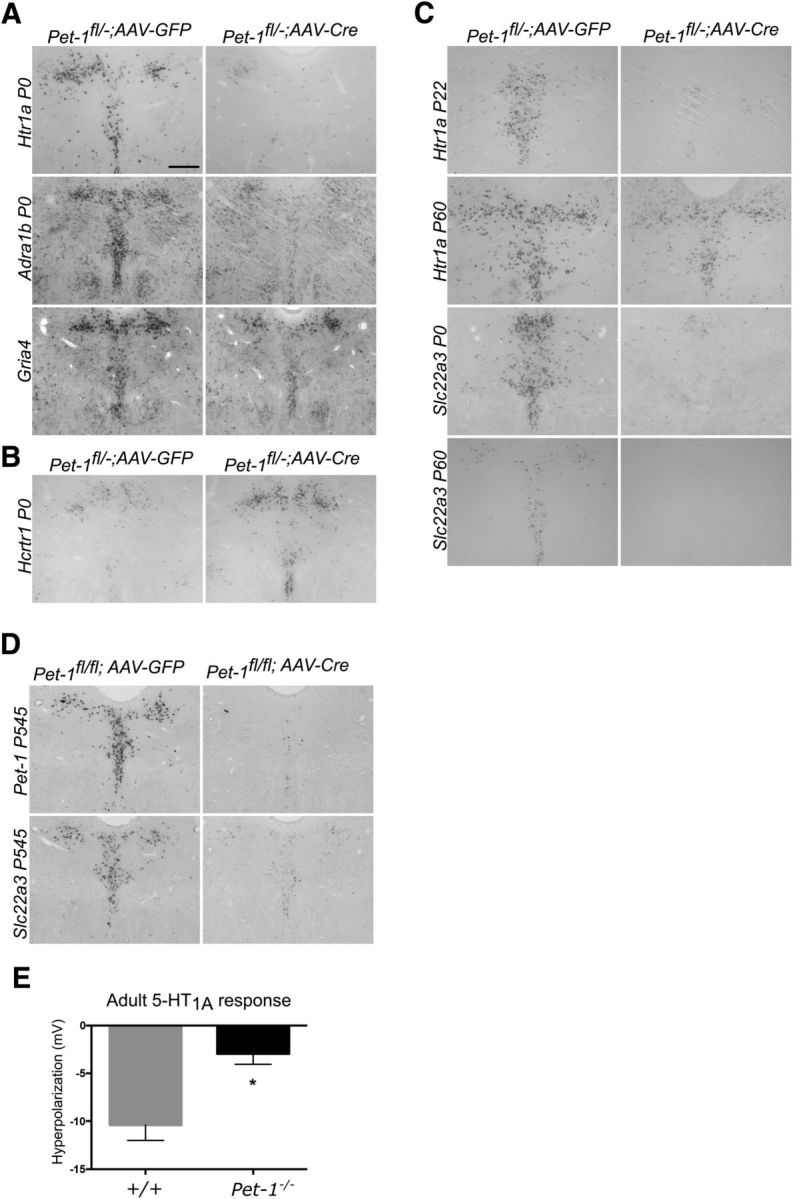
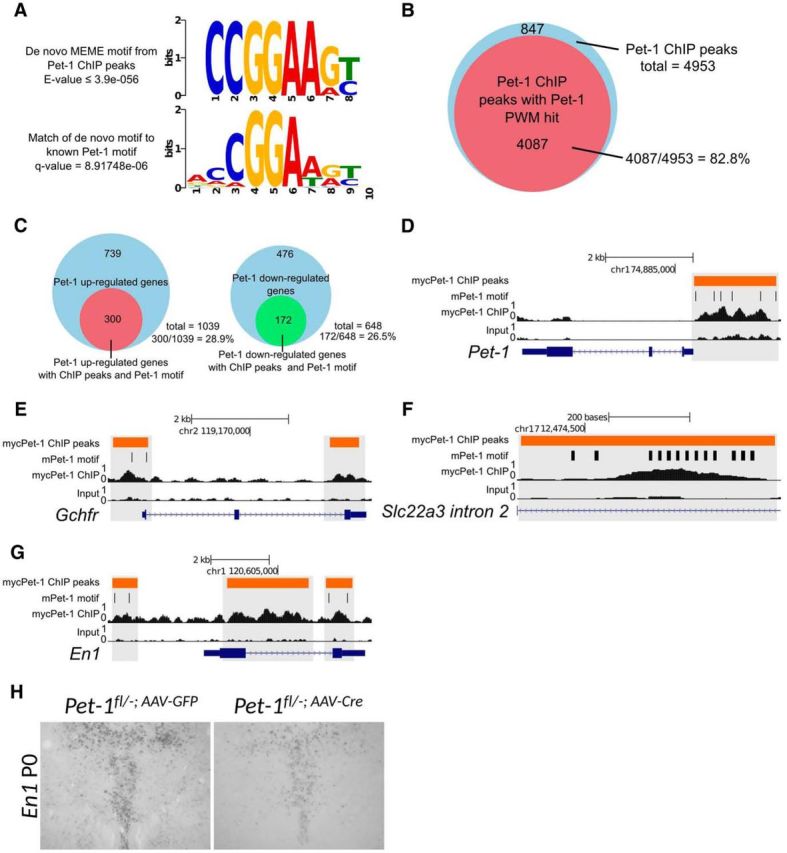
Newborn neurons enter an extended maturation stage, during which they acquire excitability characteristics crucial for development of presynaptic and postsynaptic connectivity. In contrast to earlier specification programs, little is known about the regulatory mechanisms that control neuronal maturation. The Pet-1 ETS (E26 transformation-specific) factor is continuously expressed in serotonin (5-HT) neurons and initially acts in postmitotic precursors to control acquisition of 5-HT transmitter identity. Using a combination of RNA sequencing, electrophysiology, and conditional targeting approaches, we determined gene expression patterns in maturing flow-sorted 5-HT neurons and the temporal requirements for Pet-1 in shaping these patterns for functional maturation of mouse 5-HT neurons. We report a profound disruption of postmitotic expression trajectories in Pet-1(-/-) neurons, which prevented postnatal maturation of 5-HT neuron passive and active intrinsic membrane properties, G-protein signaling, and synaptic responses to glutamatergic, lysophosphatidic, and adrenergic agonists. Unexpectedly, conditional targeting revealed a postnatal stage-specific switch in Pet-1 targets from 5-HT synthesis genes to transmitter receptor genes required for afferent modulation of 5-HT neuron excitability. Five-HT1a autoreceptor expression depended transiently on Pet-1, thus revealing an early postnatal sensitive period for control of 5-HT excitability genes. Chromatin immunoprecipitation followed by sequencing revealed that Pet-1 regulates 5-HT neuron maturation through direct gene activation and repression. Moreover, Pet-1 directly regulates the 5-HT neuron maturation factor Engrailed 1, which suggests Pet-1 orchestrates maturation through secondary postmitotic regulatory factors. The early postnatal switch in Pet-1 targets uncovers a distinct neonatal stage-specific function for Pet-1, during which it promotes maturation of 5-HT neuron excitability.
Significance statement: The regulatory mechanisms that control functional maturation of neurons are poorly understood. We show that in addition to inducing brain serotonin (5-HT) synthesis and reuptake, the Pet-1 ETS (E26 transformation-specific) factor subsequently globally coordinates postmitotic expression trajectories of genes necessary for maturation of 5-HT neuron excitability. Further, Pet-1 switches its transcriptional targets as 5-HT neurons mature from 5-HT synthesis genes to G-protein-coupled receptors, which are necessary for afferent synaptic modulation of 5-HT neuron excitability. Our findings uncover gene-specific switching of downstream targets as a previously unrecognized regulatory strategy through which continuously expressed transcription factors control acquisition of neuronal identity at different stages of development.
Keywords: Pet-1; adrenergic signaling; gene regulatory network; genomics; neuronal maturation; serotonin.
Copyright © 2016 the authors 0270-6474/16/361758-17$15.00/0.
Figures
Similar articles
-
Regulatory Mechanisms Controlling Maturation of Serotonin Neuron Identity and Function.Front Cell Neurosci. 2017 Jul 19;11:215. doi: 10.3389/fncel.2017.00215. eCollection 2017. Front Cell Neurosci. 2017. PMID: 28769770 Free PMC article.
-
Pet-1 is required across different stages of life to regulate serotonergic function.Nat Neurosci. 2010 Oct;13(10):1190-8. doi: 10.1038/nn.2623. Epub 2010 Sep 5. Nat Neurosci. 2010. PMID: 20818386 Free PMC article.
-
Pet-1 Controls Tetrahydrobiopterin Pathway and Slc22a3 Transporter Genes in Serotonin Neurons.ACS Chem Neurosci. 2015 Jul 15;6(7):1198-205. doi: 10.1021/cn500331z. Epub 2015 Feb 18. ACS Chem Neurosci. 2015. PMID: 25642596 Free PMC article.
-
Molecular genetics of mouse serotonin neurons across the lifespan.Neuroscience. 2011 Dec 1;197:17-27. doi: 10.1016/j.neuroscience.2011.08.061. Epub 2011 Aug 30. Neuroscience. 2011. PMID: 21920412 Free PMC article. Review.
-
Serotonin neuron development: shaping molecular and structural identities.Wiley Interdiscip Rev Dev Biol. 2018 Jan;7(1):10.1002/wdev.301. doi: 10.1002/wdev.301. Epub 2017 Oct 26. Wiley Interdiscip Rev Dev Biol. 2018. PMID: 29072810 Free PMC article. Review.
Cited by
-
Dorsal raphe serotonin neurotransmission is required for the expression of nursing behavior and for pup survival.Sci Rep. 2021 Mar 16;11(1):6004. doi: 10.1038/s41598-021-84368-6. Sci Rep. 2021. PMID: 33727585 Free PMC article.
-
Maintenance of neuronal identity in C. elegans and beyond: Lessons from transcription and chromatin factors.Semin Cell Dev Biol. 2024 Feb 15;154(Pt A):35-47. doi: 10.1016/j.semcdb.2023.07.001. Epub 2023 Jul 11. Semin Cell Dev Biol. 2024. PMID: 37438210 Free PMC article. Review.
-
Comparative Transcriptomic Analyses of Developing Melanocortin Neurons Reveal New Regulators for the Anorexigenic Neuron Identity.J Neurosci. 2020 Apr 15;40(16):3165-3177. doi: 10.1523/JNEUROSCI.0155-20.2020. Epub 2020 Mar 25. J Neurosci. 2020. PMID: 32213554 Free PMC article.
-
Repeated morphine exposure activates synaptogenesis and other neuroplasticity-related gene networks in the dorsomedial prefrontal cortex of male and female rats.Drug Alcohol Depend. 2021 Apr 1;221:108598. doi: 10.1016/j.drugalcdep.2021.108598. Epub 2021 Feb 13. Drug Alcohol Depend. 2021. PMID: 33626484 Free PMC article.
-
LMX1B missense-perturbation of regulatory element footprints disrupts serotonergic forebrain axon arborization.Proc Natl Acad Sci U S A. 2025 Apr 8;122(14):e2411716122. doi: 10.1073/pnas.2411716122. Epub 2025 Apr 1. Proc Natl Acad Sci U S A. 2025. PMID: 40168115
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases