

Early Somatostatin Interneuron Connectivity Mediates the Maturation of Deep Layer Cortical Circuits
- PMID: 26844832
- PMCID: PMC4861073
- DOI: 10.1016/j.neuron.2015.11.020
Early Somatostatin Interneuron Connectivity Mediates the Maturation of Deep Layer Cortical Circuits
Abstract
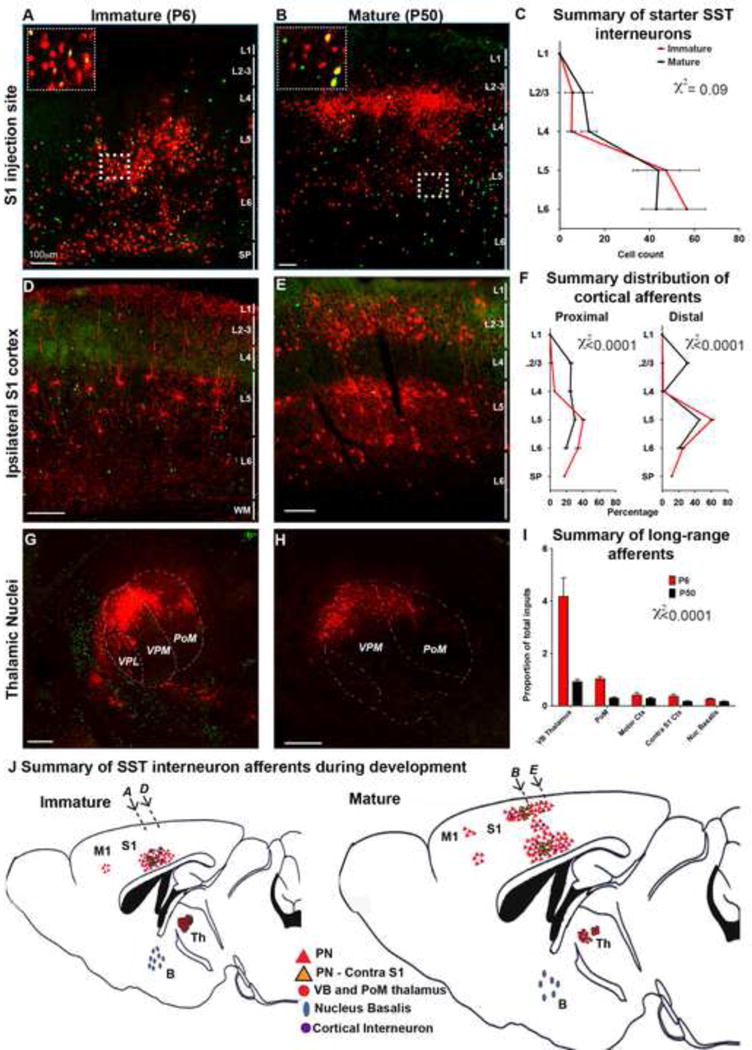
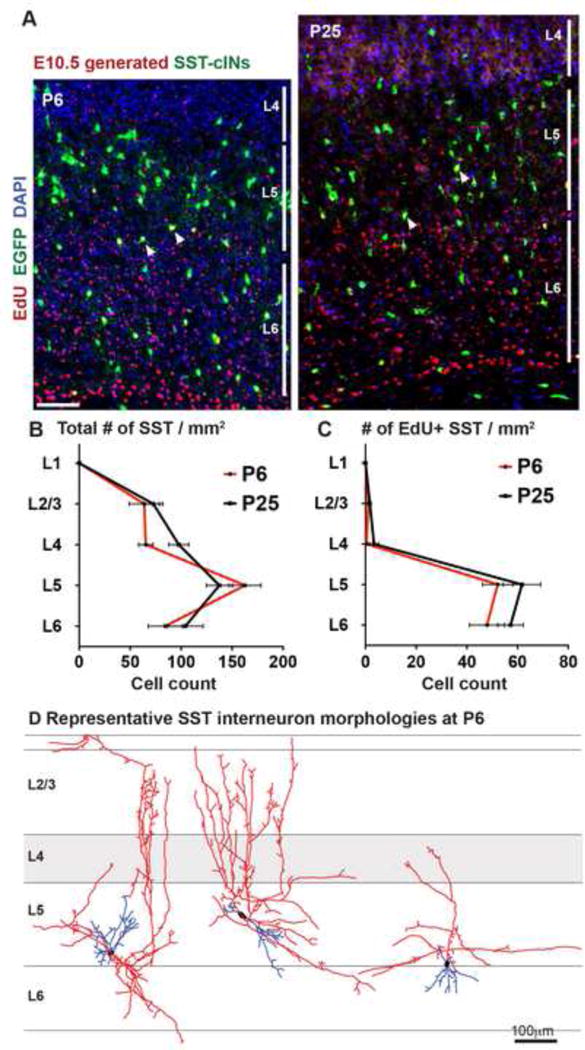
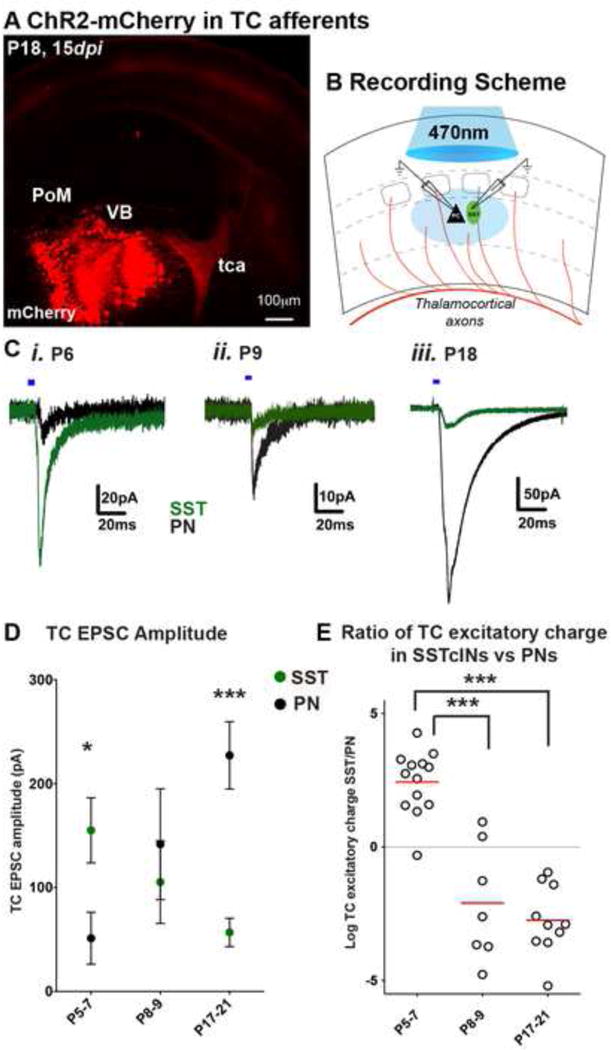
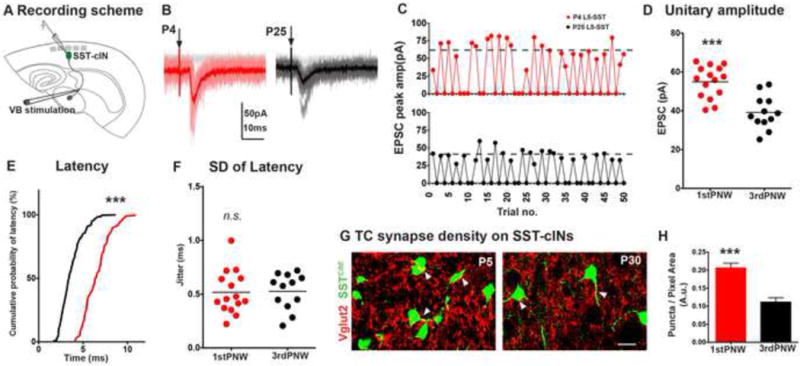
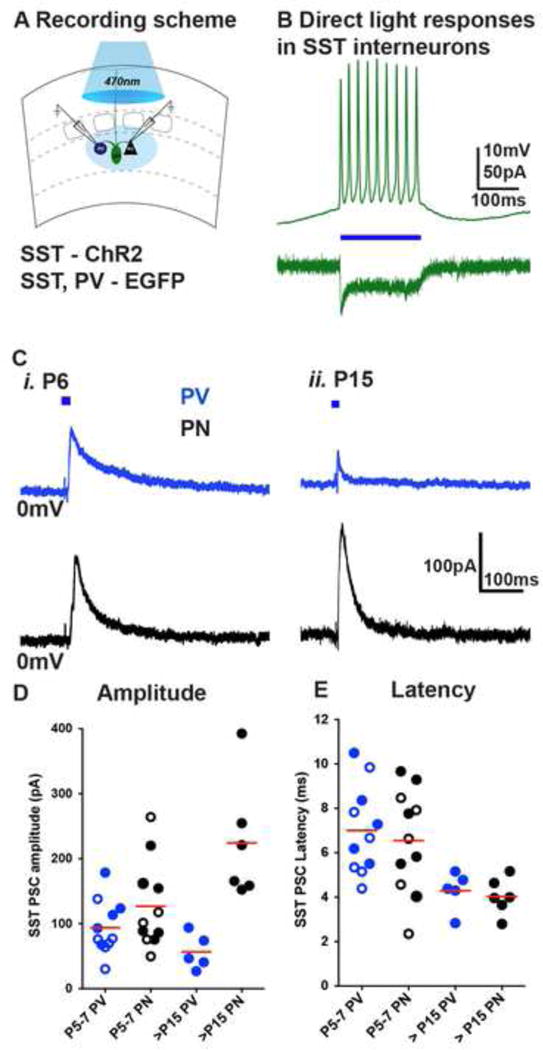
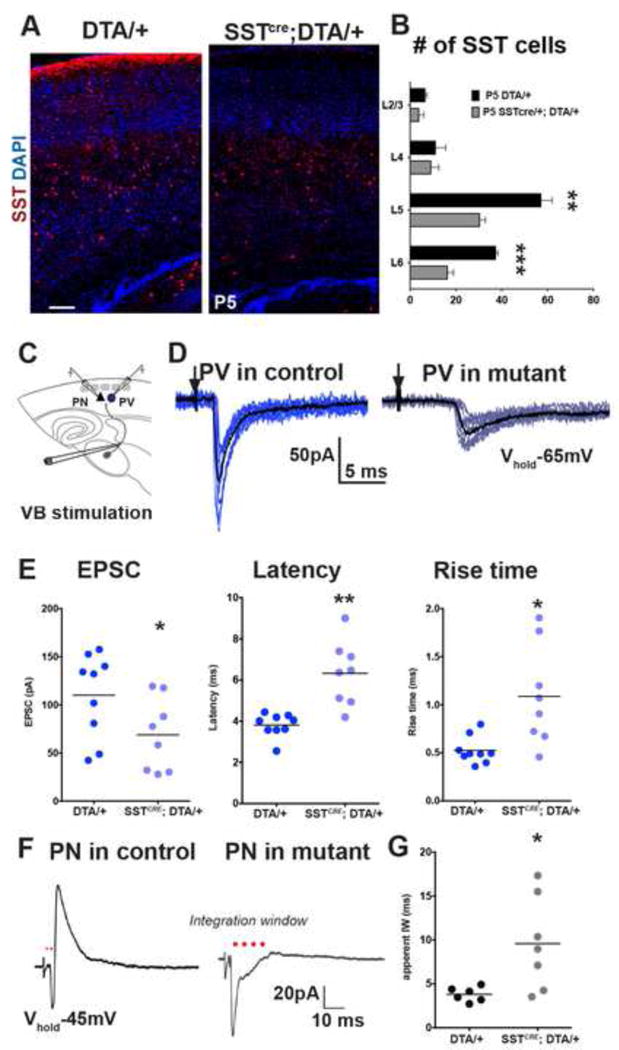
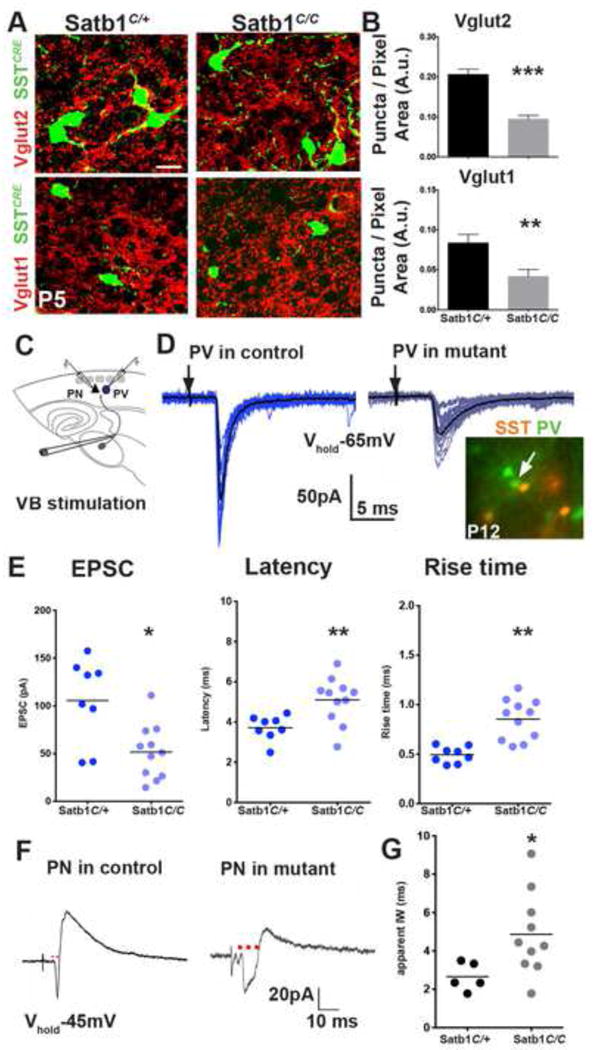
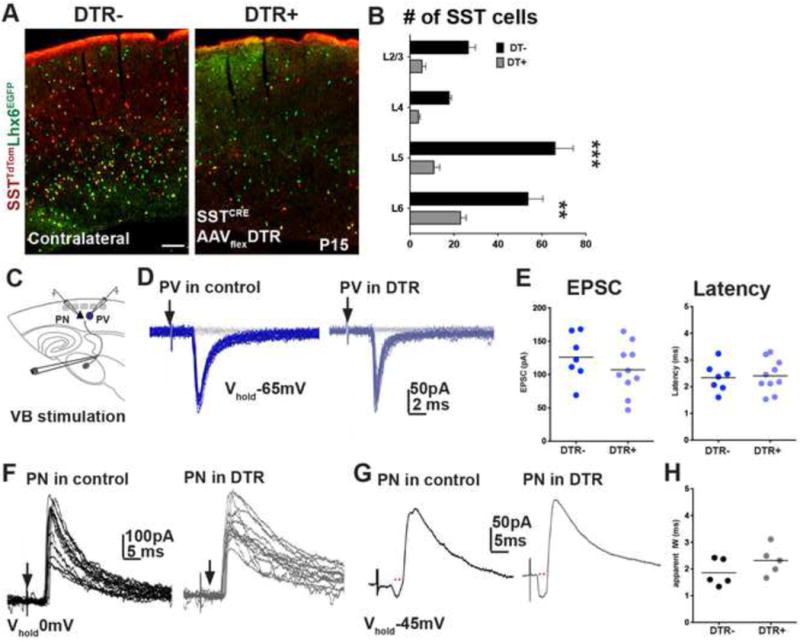
The precise connectivity of somatostatin and parvalbumin cortical interneurons is generated during development. An understanding of how these interneuron classes incorporate into cortical circuitry is incomplete but essential to elucidate the roles they play during maturation. Here, we report that somatostatin interneurons in infragranular layers receive dense but transient innervation from thalamocortical afferents during the first postnatal week. During this period, parvalbumin interneurons and pyramidal neurons within the same layers receive weaker thalamocortical inputs, yet are strongly innervated by somatostatin interneurons. Further, upon disruption of the early (but not late) somatostatin interneuron network, the synaptic maturation of thalamocortical inputs onto parvalbumin interneurons is perturbed. These results suggest that infragranular somatostatin interneurons exhibit a transient early synaptic connectivity that is essential for the establishment of thalamic feedforward inhibition mediated by parvalbumin interneurons.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Angevine JB, Jr, Sidman RL. Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature. 1961;192:766–768. - PubMed
-
- Bartolini G, Ciceri G, Marin O. Integration of GABAergic interneurons into cortical cell assemblies: lessons from embryos and adults. Neuron. 2013;79:849–864. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 NS074972/NS/NINDS NIH HHS/United States
- R01NS30989/NS/NINDS NIH HHS/United States
- R01 EY022577/EY/NEI NIH HHS/United States
- MH0639123/MH/NIMH NIH HHS/United States
- T32 NS086750/NS/NINDS NIH HHS/United States
- MH095147/MH/NIMH NIH HHS/United States
- P01NS074972/NS/NINDS NIH HHS/United States
- EY022577/EY/NEI NIH HHS/United States
- R01 MH068469/MH/NIMH NIH HHS/United States
- R01 NS086372/NS/NINDS NIH HHS/United States
- NS080586/NS/NINDS NIH HHS/United States
- NS086372/NS/NINDS NIH HHS/United States
- R01 NS080586/NS/NINDS NIH HHS/United States
- R01 MH063912/MH/NIMH NIH HHS/United States
- R01 MH095147/MH/NIMH NIH HHS/United States
- R37 MH071679/MH/NIMH NIH HHS/United States
- R01 NS081297/NS/NINDS NIH HHS/United States
- T32 MH015174/MH/NIMH NIH HHS/United States
- R01 MH071679/MH/NIMH NIH HHS/United States
- NS090919/NS/NINDS NIH HHS/United States
- NS 081297/NS/NINDS NIH HHS/United States
- R01 NS030989/NS/NINDS NIH HHS/United States
- R01 NS090919/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
