

Macrophage IKKα Deficiency Suppresses Akt Phosphorylation, Reduces Cell Survival, and Decreases Early Atherosclerosis
- PMID: 26848161
- PMCID: PMC4808396
- DOI: 10.1161/ATVBAHA.115.306931
Macrophage IKKα Deficiency Suppresses Akt Phosphorylation, Reduces Cell Survival, and Decreases Early Atherosclerosis
Abstract
Objective: The IκB kinase (IKK) is an enzyme complex that initiates the nuclear factor κB transcription factor cascade, which is important in regulating multiple cellular responses. IKKα is directly associated with 2 major prosurvival pathways, PI3K/Akt and nuclear factor κB, but its role in cell survival is not clear. Macrophages play critical roles in the pathogenesis of atherosclerosis, yet the impact of IKKα signaling on macrophage survival and atherogenesis remains unclear.
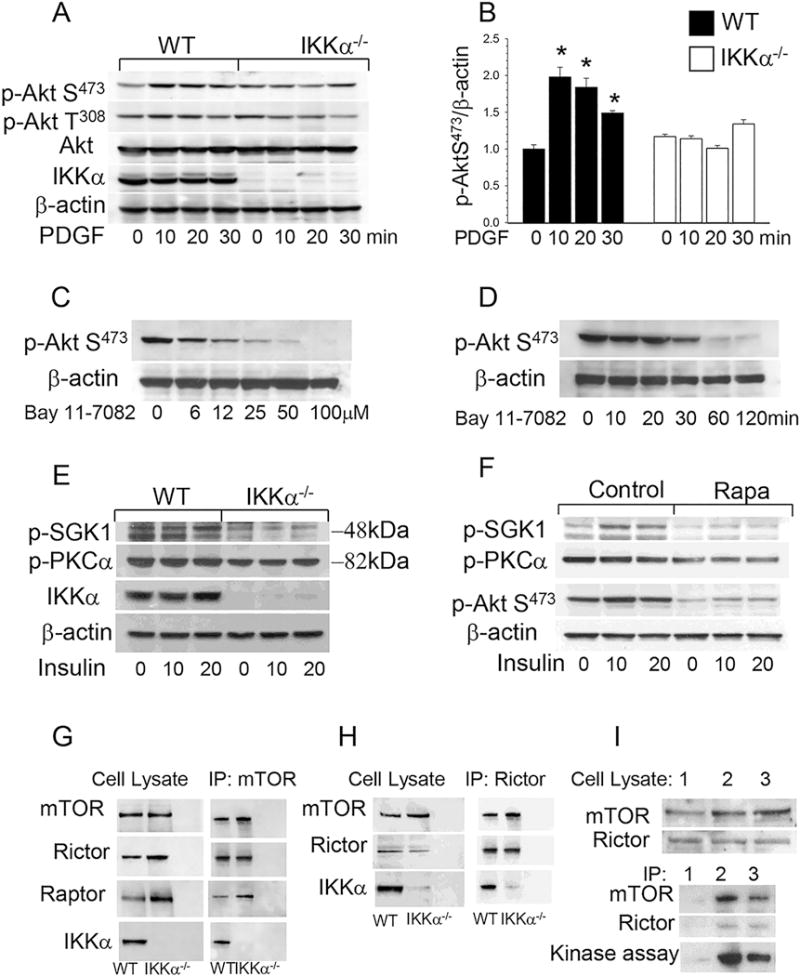
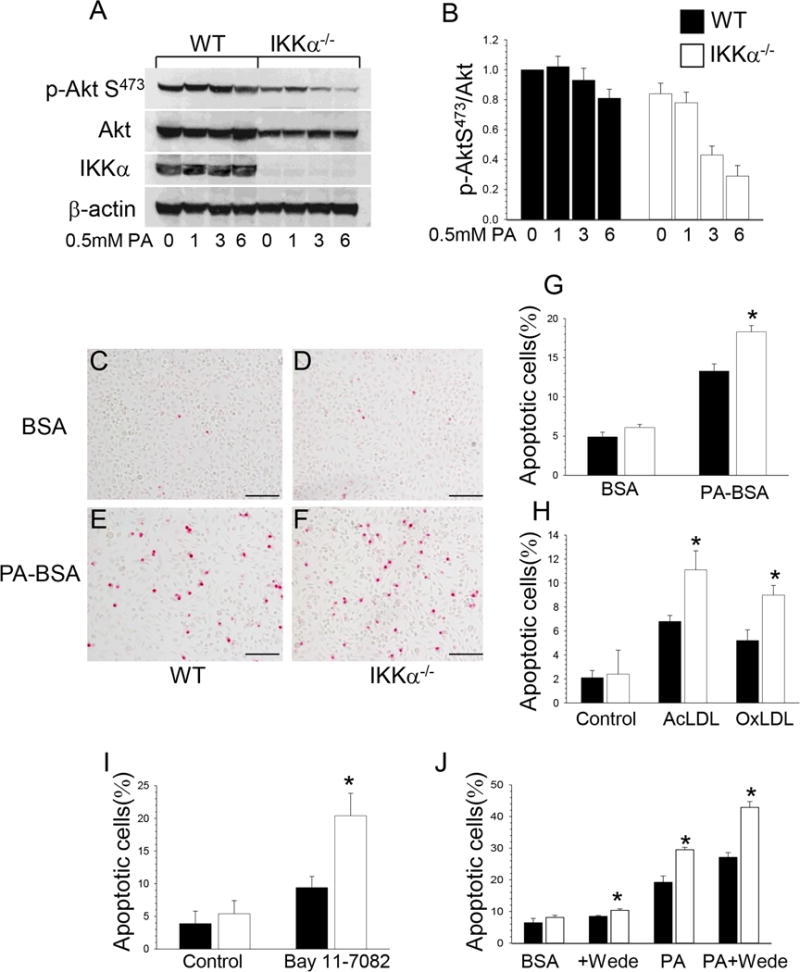
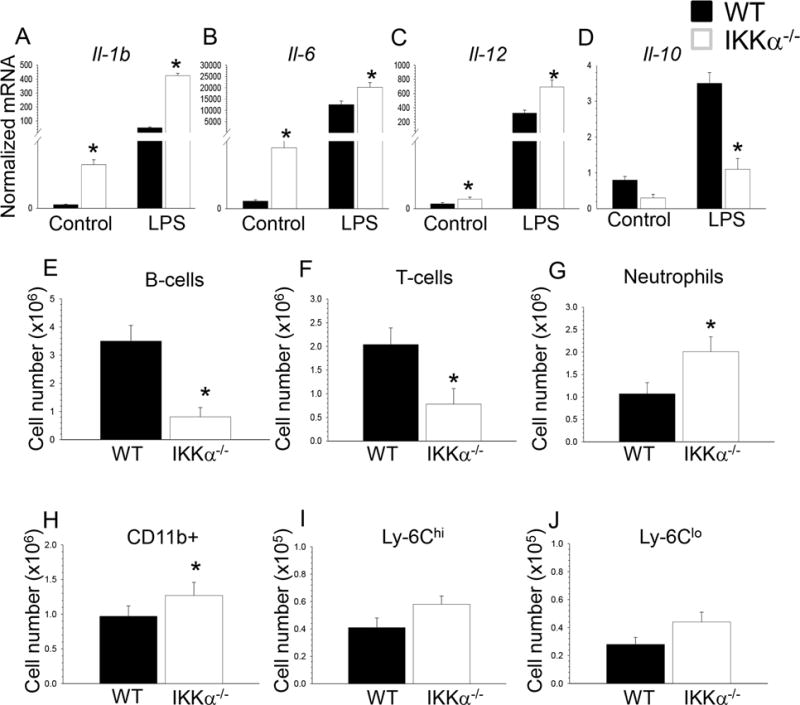
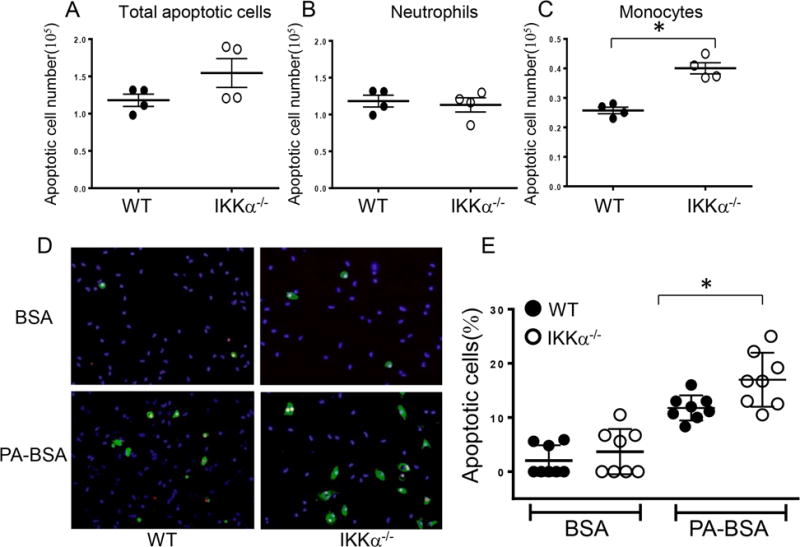
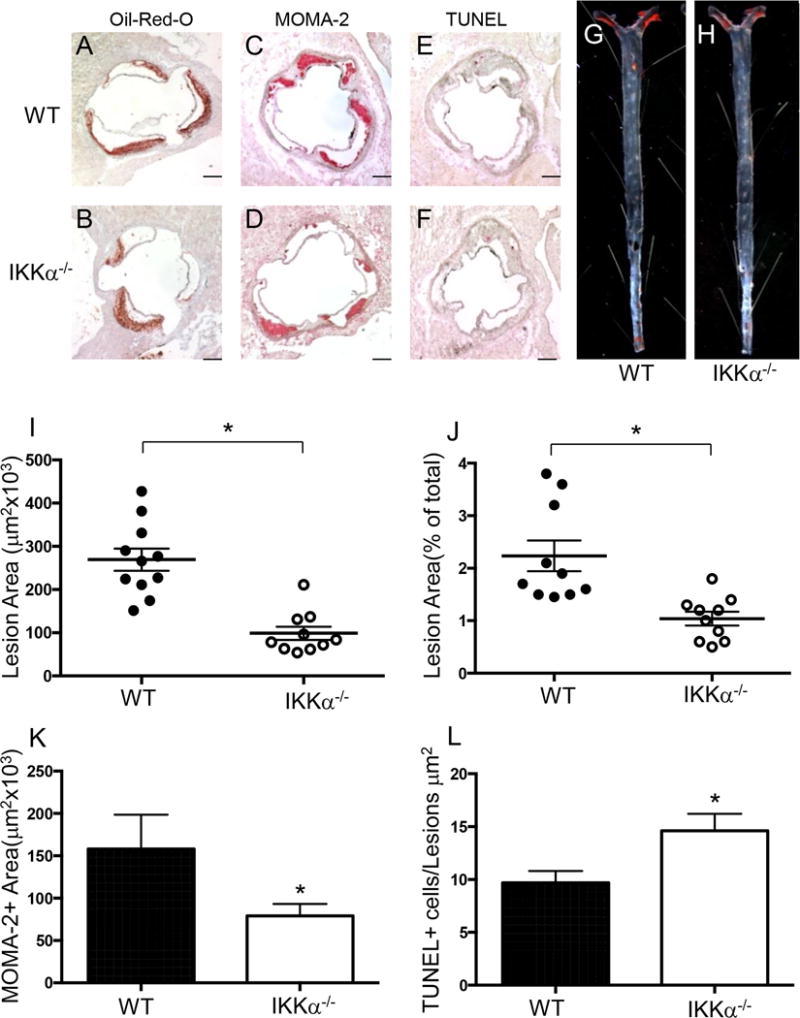
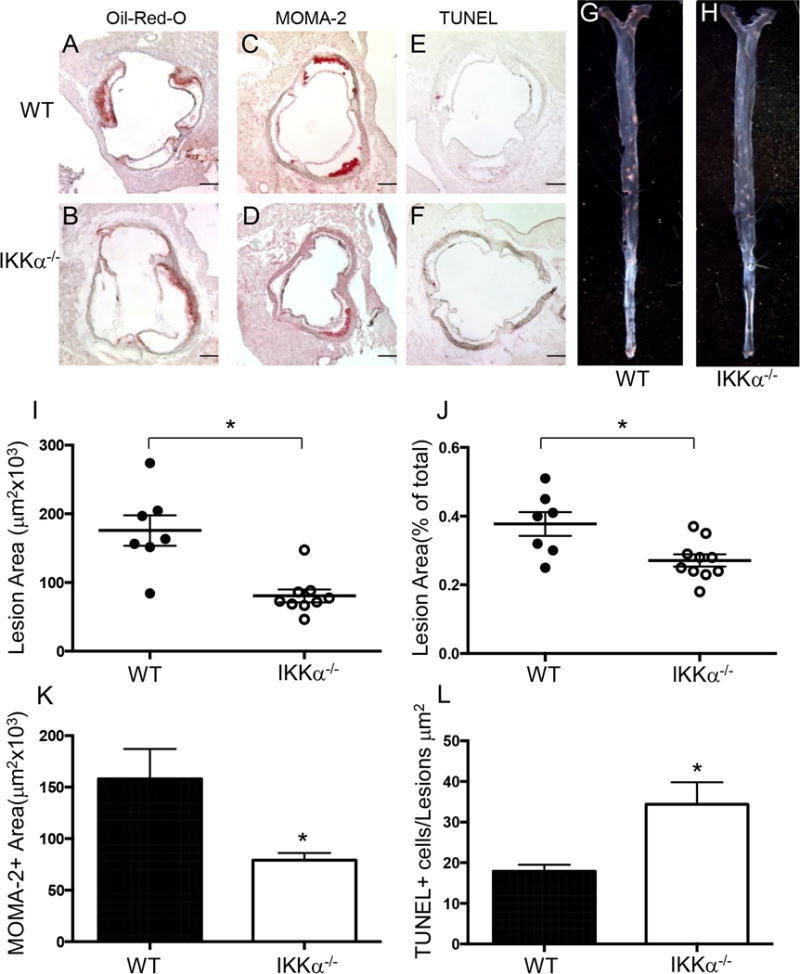
Approach and results: Here, we demonstrate that genetic IKKα deficiency, as well as pharmacological inhibition of IKK, in mouse macrophages significantly reduces Akt S(473) phosphorylation, which is accompanied by suppression of mTOR complex 2 signaling. Moreover, IKKα null macrophages treated with lipotoxic palmitic acid exhibited early exhaustion of Akt signaling compared with wild-type cells. This was accompanied by a dramatic decrease in the resistance of IKKα(-/-) monocytes and macrophages to different proapoptotic stimuli compared with wild-type cells. In vivo, IKKα deficiency increased macrophage apoptosis in atherosclerotic lesions and decreased early atherosclerosis in both female and male low-density lipoprotein receptor (LDLR)(-/-) mice reconstituted with IKKα(-/-) hematopoietic cells and fed with the Western diet for 8 weeks compared with control LDLR(-/-) mice transplanted with wild-type cells.
Conclusions: Hematopoietic IKKα deficiency in mouse suppresses Akt signaling, compromising monocyte/macrophage survival and this decreases early atherosclerosis.
Keywords: apoptosis; atherosclerosis; cell survival; macrophages; phosphorylation.
© 2016 American Heart Association, Inc.
Conflict of interest statement
Figures
References
-
- Hansson GK, Libby P. The immune response in atherosclerosis: A double-edged sword. Nat Rev Immunol. 2006;6:508–519. - PubMed
-
- Kockx MM, Herman AG. Apoptosis in atherosclerosis: Beneficial or detrimental? Cardiovasc Res. 2000;45:736–746. - PubMed
-
- Duronio V. The life of a cell: Apoptosis regulation by the pi3k/pkb pathway. Biochem J. 2008;415:333–344. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
