

Mapping and Exploring the Collagen-I Proteostasis Network
- PMID: 26848503
- PMCID: PMC4910512
- DOI: 10.1021/acschembio.5b01083
Mapping and Exploring the Collagen-I Proteostasis Network
Abstract
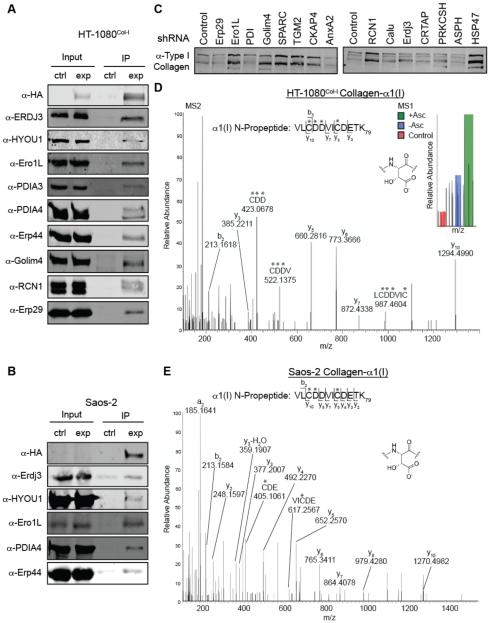
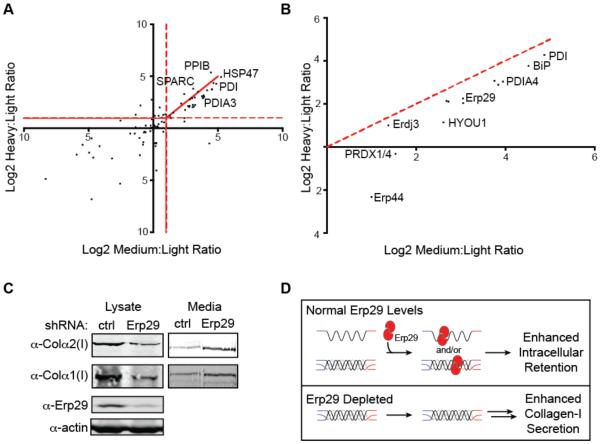
Collagen-I is the most abundant protein in the human body, yet our understanding of how the endoplasmic reticulum regulates collagen-I proteostasis (folding, quality control, and secretion) remains immature. Of particular importance, interactomic studies to map the collagen-I proteostasis network have never been performed. Such studies would provide insight into mechanisms of collagen-I folding and misfolding in cells, an area that is particularly important owing to the prominence of the collagen misfolding-related diseases. Here, we overcome key roadblocks to progress in this area by generating stable fibrosarcoma cells that inducibly express properly folded and modified collagen-I strands tagged with distinctive antibody epitopes. Selective immunoprecipitation of collagen-I from these cells integrated with quantitative mass spectrometry-based proteomics permits the first mapping of the collagen-I proteostasis network. Biochemical validation of the resulting map leads to the assignment of numerous new players in collagen-I proteostasis, and the unanticipated discovery of apparent aspartyl-hydroxylation as a new post-translational modification in the N-propeptide of collagen-I. Furthermore, quantitative analyses reveal that Erp29, an abundant endoplasmic reticulum proteostasis machinery component with few known functions, plays a key role in collagen-I retention under ascorbate-deficient conditions. In summary, the work here provides fresh insights into the molecular mechanisms of collagen-I proteostasis, yielding a detailed roadmap for future investigations. Straightforward adaptations of the cellular platform developed will also enable hypothesis-driven, comparative research on the likely distinctive proteostasis mechanisms engaged by normal and disease-causing, misfolding collagen-I variants, potentially motivating new therapeutic strategies for currently incurable collagenopathies.
Figures
Similar articles
-
Elucidation of proteostasis defects caused by osteogenesis imperfecta mutations in the collagen-α2(I) C-propeptide domain.J Biol Chem. 2020 Jul 17;295(29):9959-9973. doi: 10.1074/jbc.RA120.014071. Epub 2020 Jun 1. J Biol Chem. 2020. PMID: 32482890 Free PMC article.
-
Targeting defective proteostasis in the collagenopathies.Curr Opin Chem Biol. 2019 Jun;50:80-88. doi: 10.1016/j.cbpa.2019.02.021. Epub 2019 Apr 24. Curr Opin Chem Biol. 2019. PMID: 31028939 Free PMC article. Review.
-
Mass Spectrometry-Based Proteomics to Define Intracellular Collagen Interactomes.Methods Mol Biol. 2019;1944:95-114. doi: 10.1007/978-1-4939-9095-5_7. Methods Mol Biol. 2019. PMID: 30840237 Free PMC article.
-
Revealing functional insights into ER proteostasis through proteomics and interactomics.Exp Cell Res. 2021 Feb 1;399(1):112417. doi: 10.1016/j.yexcr.2020.112417. Epub 2020 Dec 8. Exp Cell Res. 2021. PMID: 33301765 Free PMC article. Review.
-
Proteostasis: bad news and good news from the endoplasmic reticulum.Swiss Med Wkly. 2014 Aug 21;144:w14001. doi: 10.4414/smw.2014.14001. eCollection 2014. Swiss Med Wkly. 2014. PMID: 25144910 Review.
Cited by
-
Collagen's enigmatic, highly conserved N-glycan has an essential proteostatic function.Proc Natl Acad Sci U S A. 2021 Mar 9;118(10):e2026608118. doi: 10.1073/pnas.2026608118. Proc Natl Acad Sci U S A. 2021. PMID: 33674390 Free PMC article.
-
The Probable, Possible, and Novel Functions of ERp29.Front Physiol. 2020 Sep 8;11:574339. doi: 10.3389/fphys.2020.574339. eCollection 2020. Front Physiol. 2020. PMID: 33013490 Free PMC article.
-
LOX-catalyzed collagen stabilization is a proximal cause for intrinsic resistance to chemotherapy.Oncogene. 2018 Sep;37(36):4921-4940. doi: 10.1038/s41388-018-0320-2. Epub 2018 May 21. Oncogene. 2018. PMID: 29780168 Free PMC article.
-
Adapting Secretory Proteostasis and Function Through the Unfolded Protein Response.Curr Top Microbiol Immunol. 2018;414:1-25. doi: 10.1007/82_2017_56. Curr Top Microbiol Immunol. 2018. PMID: 28929194 Free PMC article. Review.
-
A cysteine-based molecular code informs collagen C-propeptide assembly.Nat Commun. 2018 Oct 11;9(1):4206. doi: 10.1038/s41467-018-06185-2. Nat Commun. 2018. PMID: 30310058 Free PMC article.
References
-
- Ishikawa Y, Bächinger HP. A molecular ensemble in the rER for procollagen maturation. Biochim. Biophys. Acta. 2013;1833:2479–2491. - PubMed
-
- Myllyharju J, Kivirikko KI. Collagens and collagen-related diseases. Ann. Med. 2001;33:7–21. - PubMed
-
- Tosi LL, Warman ML. Mechanistic and therapeutic insights gained from studying rare skeletal diseases. Bone. 2015;76:67–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
