

Structural Insights into Mycobacterium tuberculosis Rv2671 Protein as a Dihydrofolate Reductase Functional Analogue Contributing to para-Aminosalicylic Acid Resistance
- PMID: 26848874
- PMCID: PMC6201685
- DOI: 10.1021/acs.biochem.5b00993
Structural Insights into Mycobacterium tuberculosis Rv2671 Protein as a Dihydrofolate Reductase Functional Analogue Contributing to para-Aminosalicylic Acid Resistance
Abstract
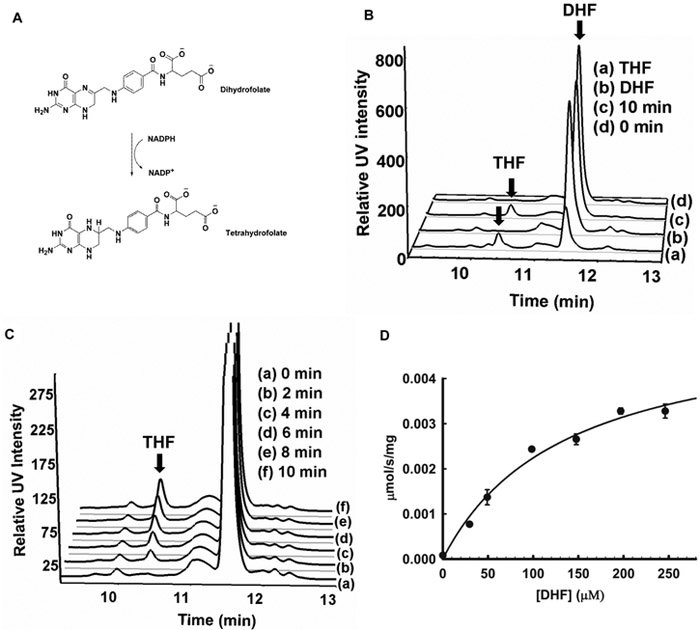
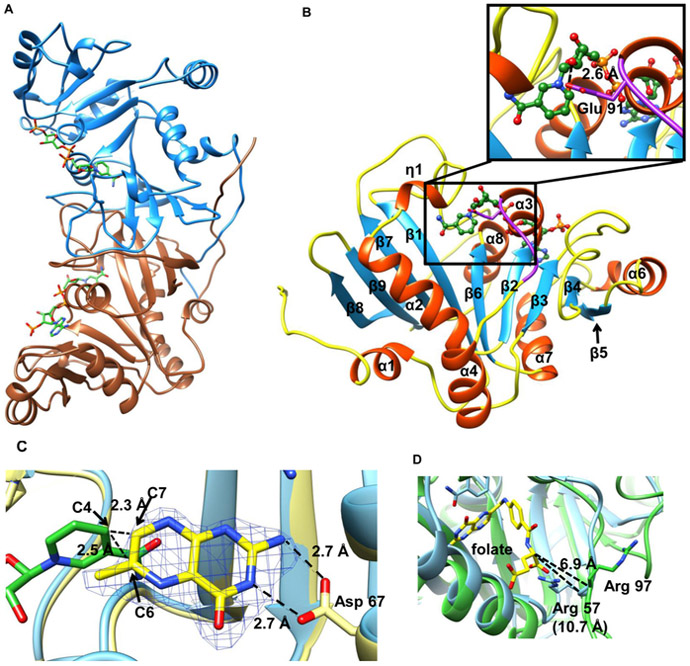
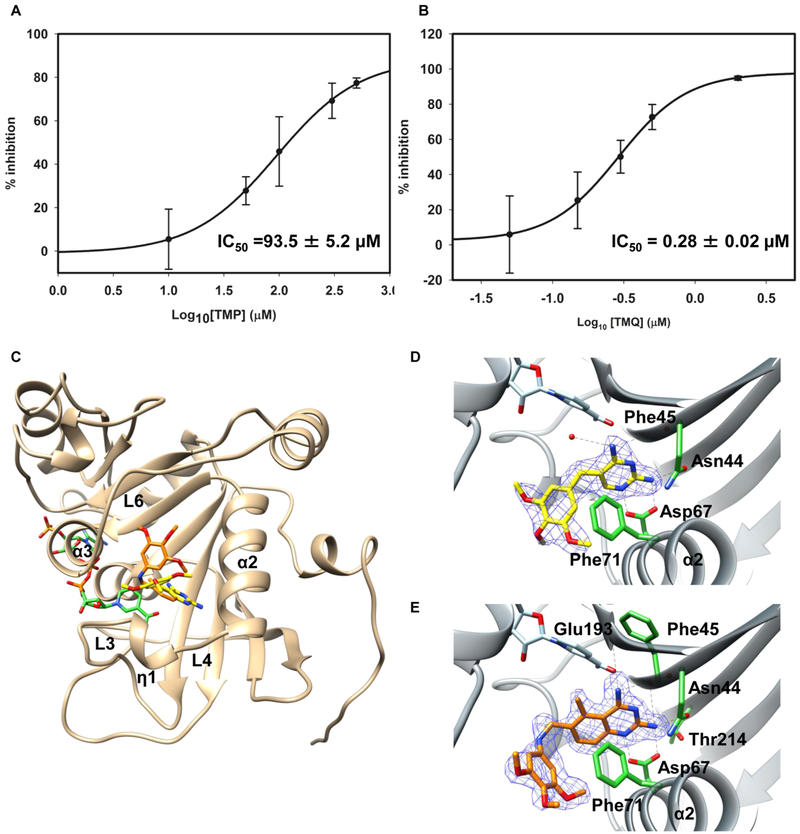
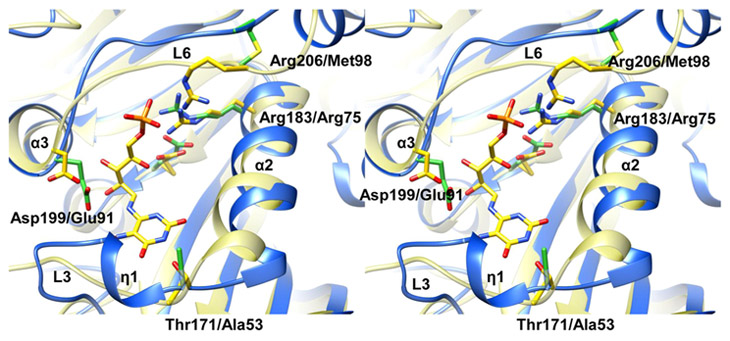
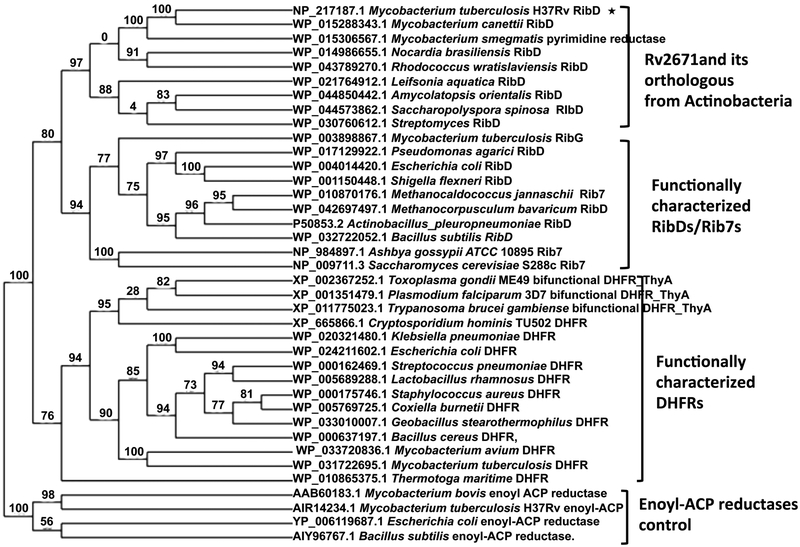
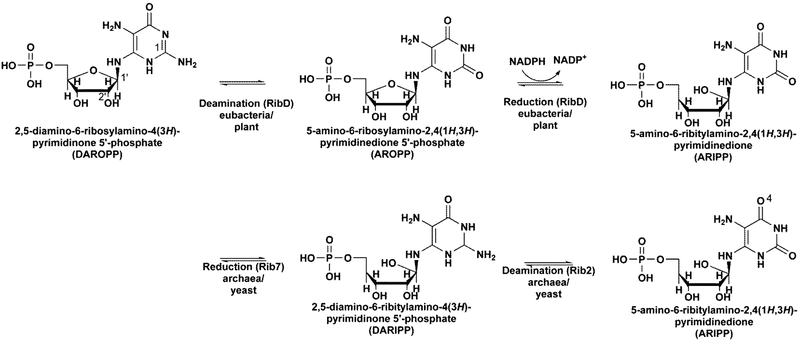
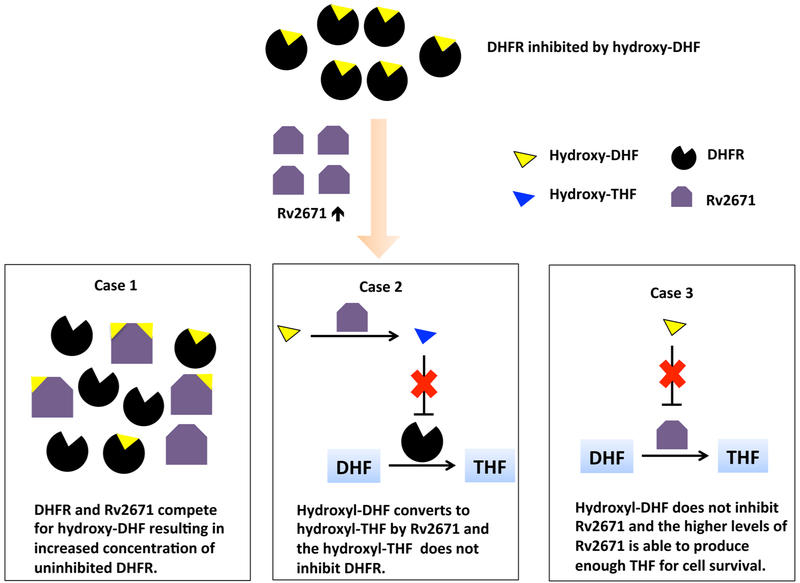
Mycobacterium tuberculosis (Mtb) Rv2671 is annotated as a 5-amino-6-ribitylamino-2,4(1H,3H)-pyrimidinedione 5'-phosphate (AROPP) reductase (RibD) in the riboflavin biosynthetic pathway. Recently, a strain of Mtb with a mutation in the 5' untranslated region of Rv2671, which resulted in its overexpression, was found to be resistant to dihydrofolate reductase (DHFR) inhibitors including the anti-Mtb drug para-aminosalicylic acid (PAS). In this study, a biochemical analysis of Rv2671 showed that it was able to catalyze the reduction of dihydrofolate (DHF) to tetrahydrofolate (THF), which explained why the overexpression of Rv2671 was sufficient to confer PAS resistance. We solved the structure of Rv2671 in complex with the NADP(+) and tetrahydrofolate (THF), which revealed the structural basis for the DHFR activity. The structures of Rv2671 complexed with two DHFR inhibitors, trimethoprim and trimetrexate, provided additional details of the substrate binding pocket and elucidated the differences between their inhibitory activities. Finally, Rv2671 was unable to catalyze the reduction of AROPP, which indicated that Rv2671 and its closely related orthologues are not involved in riboflavin biosynthesis.
Figures
References
-
- CDC. Update: Availability of Streptomycin and para-Amino-salicylic Acid—United States, Morbidity and Mortality Weekly Report; 41, 26; Center for Disease Control and Prevention: Atlanta, GA, 1992; 482.
-
- Rieckmann KH (1973) The in Vitro Activity of Experimental Antimalarial Compounds against Strains of Plasmodium falciparum with Varying Degrees of Sensitivity to Pyrimethamine and Chloroquine, in Chemotherapy of Malaria and Resistance to Antimalarials, 529th ed., p 58, WHO Tech. Rep. Ser., World Health Organization, Geneva, Switzerland.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
