

Voltage-Gated K+ Channel, Kv3.3 Is Involved in Hemin-Induced K562 Differentiation
- PMID: 26849432
- PMCID: PMC4743930
- DOI: 10.1371/journal.pone.0148633
Voltage-Gated K+ Channel, Kv3.3 Is Involved in Hemin-Induced K562 Differentiation
Abstract
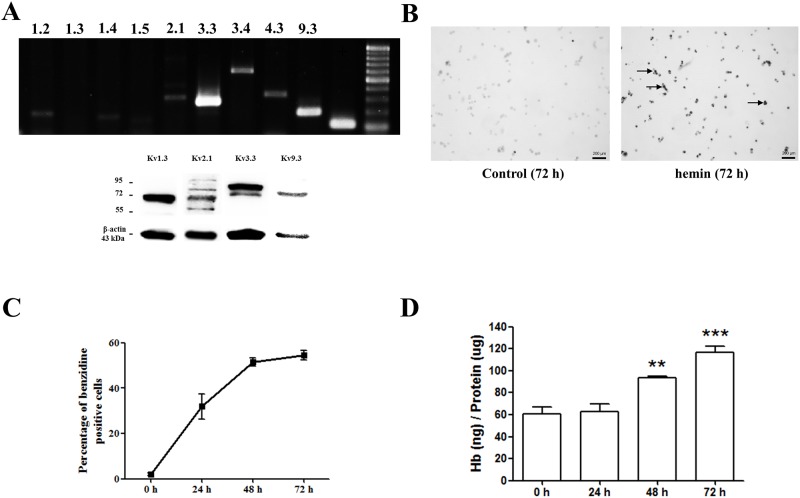
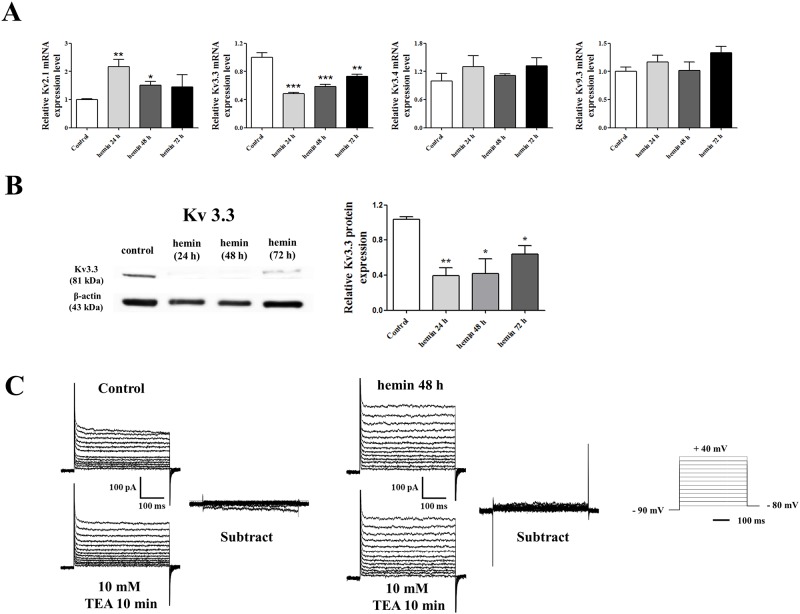
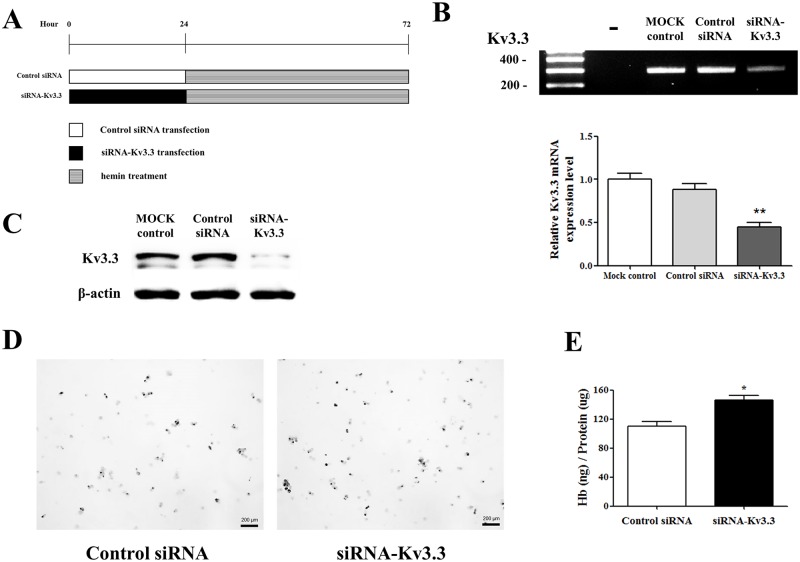
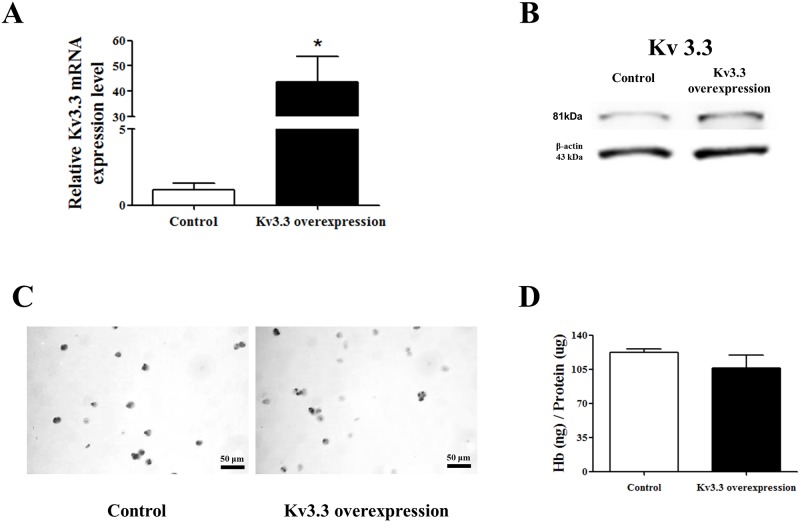
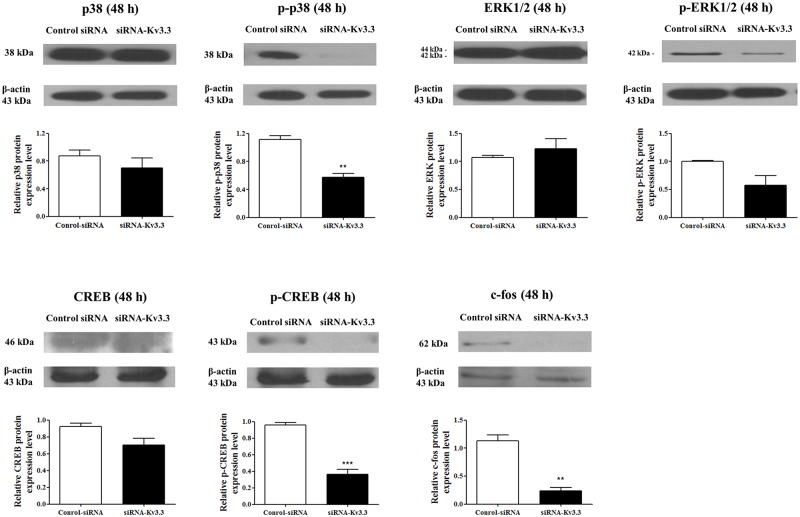
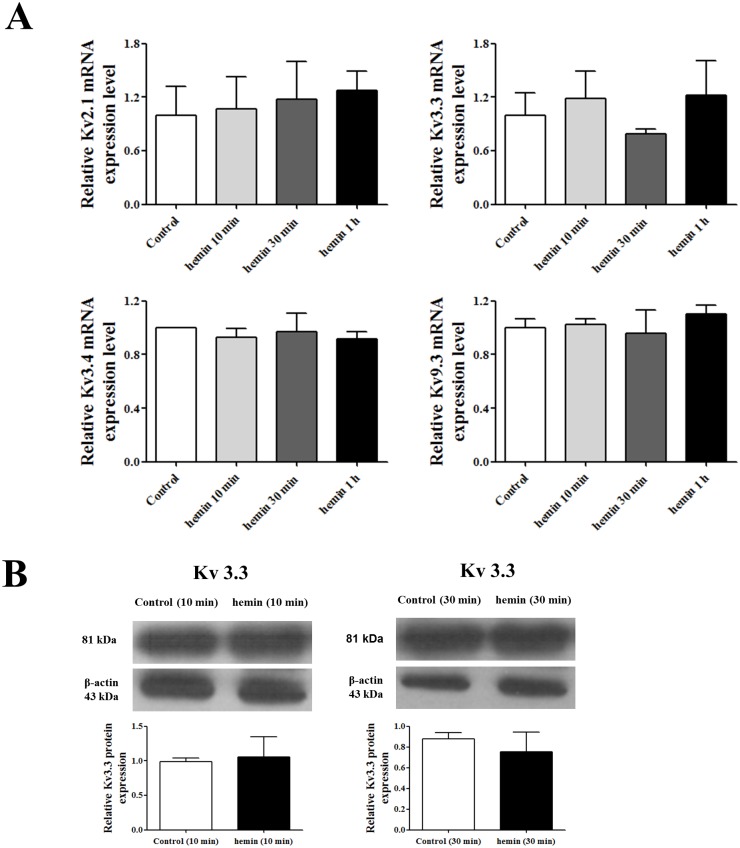
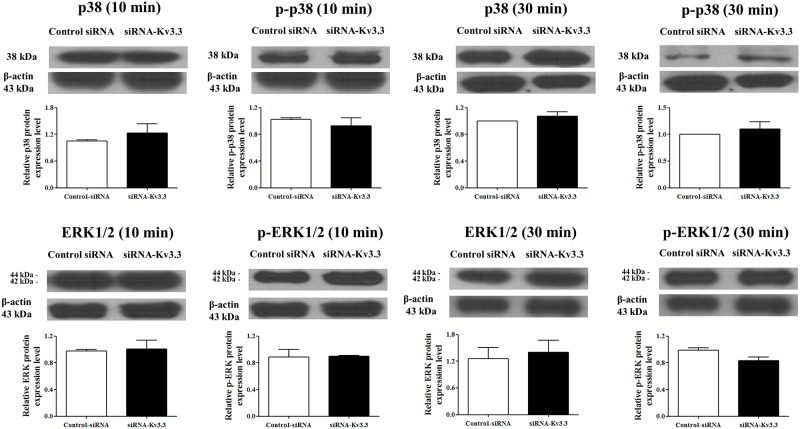
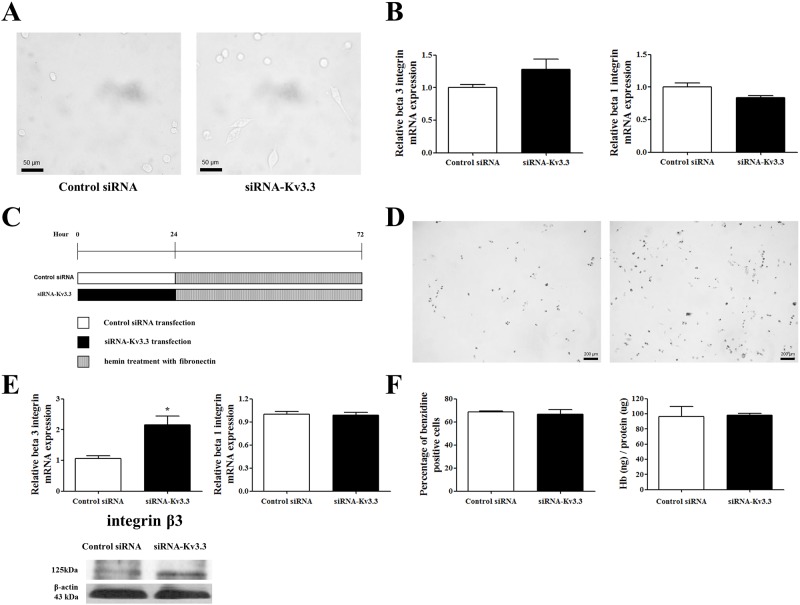
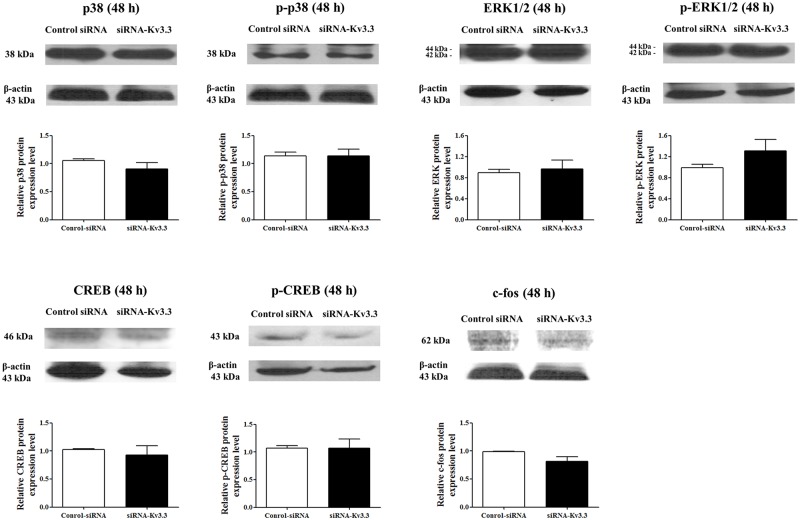
Voltage-gated K+ (Kv) channels are well known to be involved in cell proliferation. However, even though cell proliferation is closely related to cell differentiation, the relationship between Kv channels and cell differentiation remains poorly investigated. This study demonstrates that Kv3.3 is involved in K562 cell erythroid differentiation. Down-regulation of Kv3.3 using siRNA-Kv3.3 increased hemin-induced K562 erythroid differentiation through decreased activation of signal molecules such as p38, cAMP response element-binding protein, and c-fos. Down-regulation of Kv3.3 also enhanced cell adhesion by increasing integrin β3 and this effect was amplified when the cells were cultured with fibronectin. The Kv channels, or at least Kv3.3, appear to be associated with cell differentiation; therefore, understanding the mechanisms of Kv channel regulation of cell differentiation would provide important information regarding vital cellular processes.
Conflict of interest statement
Figures
References
-
- Hille B. Ion channels of excitable membranes. 3rd ed Sunderland: Sinauer associates, Inc; 2001.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
