

Structural Basis and Functional Role of Intramembrane Trimerization of the Fas/CD95 Death Receptor
- PMID: 26853147
- PMCID: PMC4761300
- DOI: 10.1016/j.molcel.2016.01.009
Structural Basis and Functional Role of Intramembrane Trimerization of the Fas/CD95 Death Receptor
Abstract
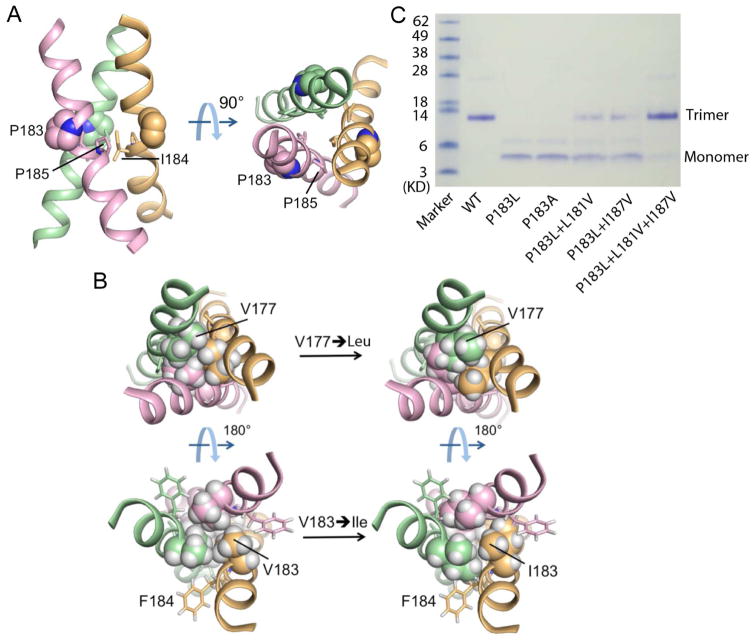
Fas (CD95, Apo-1, or TNFRSF6) is a prototypical apoptosis-inducing death receptor in the tumor necrosis factor receptor (TNFR) superfamily. While the extracellular domains of TNFRs form trimeric complexes with their ligands and the intracellular domains engage in higher-order oligomerization, the role of the transmembrane (TM) domains is unknown. We determined the NMR structures of mouse and human Fas TM domains in bicelles that mimic lipid bilayers. Surprisingly, these domains use proline motifs to create optimal packing in homotrimer assembly distinct from classical trimeric coiled-coils in solution. Cancer-associated and structure-based mutations in Fas TM disrupt trimerization in vitro and reduce apoptosis induction in vivo, indicating the essential role of intramembrane trimerization in receptor activity. Our data suggest that the structures represent the signaling-active conformation of Fas TM, which appears to be different from the pre-ligand conformation. Analysis of other TNFR sequences suggests proline-containing sequences as common motifs for receptor TM trimerization.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
