

Aging augments the impact of influenza respiratory tract infection on mobility impairments, muscle-localized inflammation, and muscle atrophy
- PMID: 26856410
- PMCID: PMC4925818
- DOI: 10.18632/aging.100882
Aging augments the impact of influenza respiratory tract infection on mobility impairments, muscle-localized inflammation, and muscle atrophy
Abstract
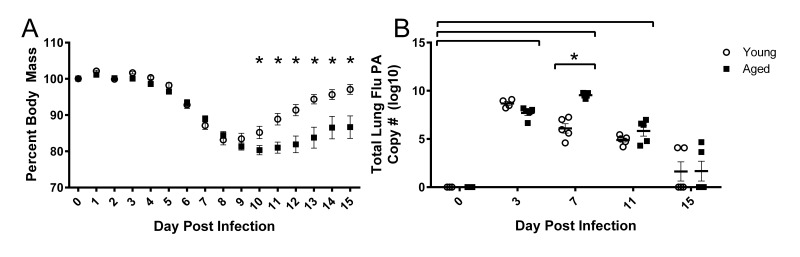
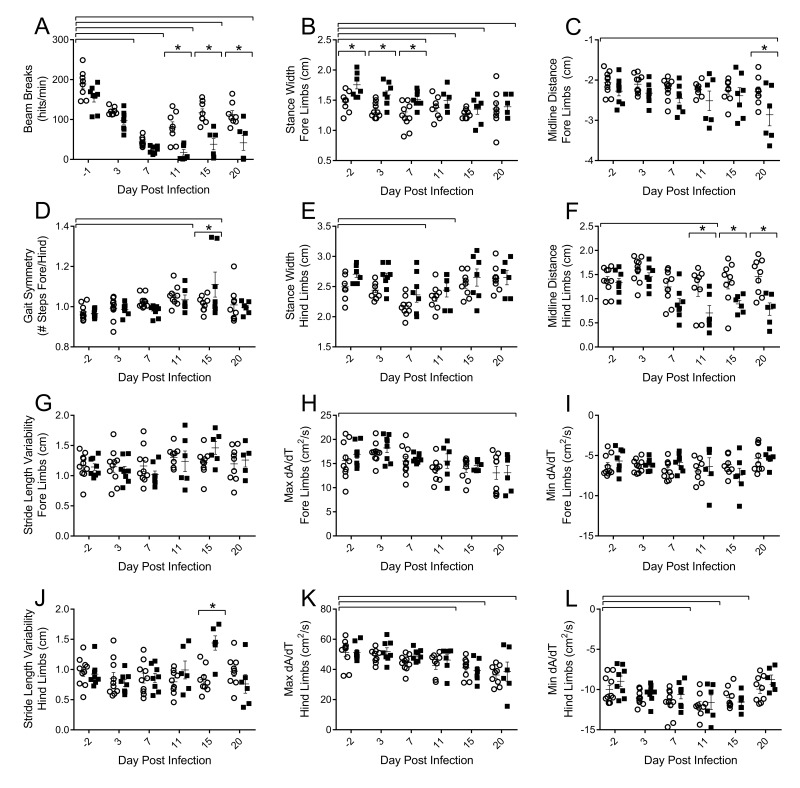
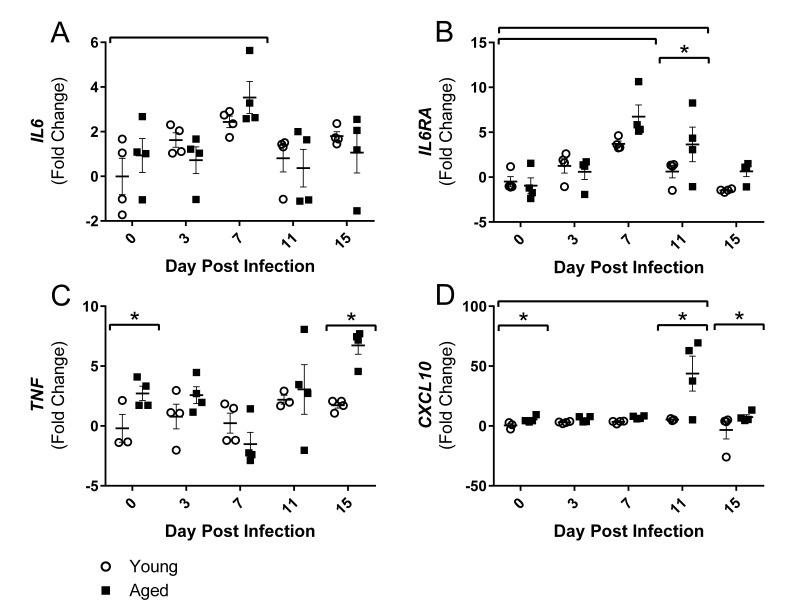
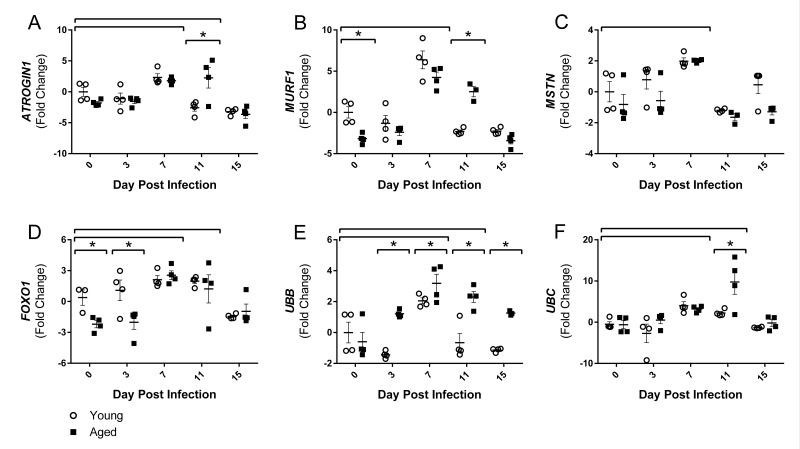
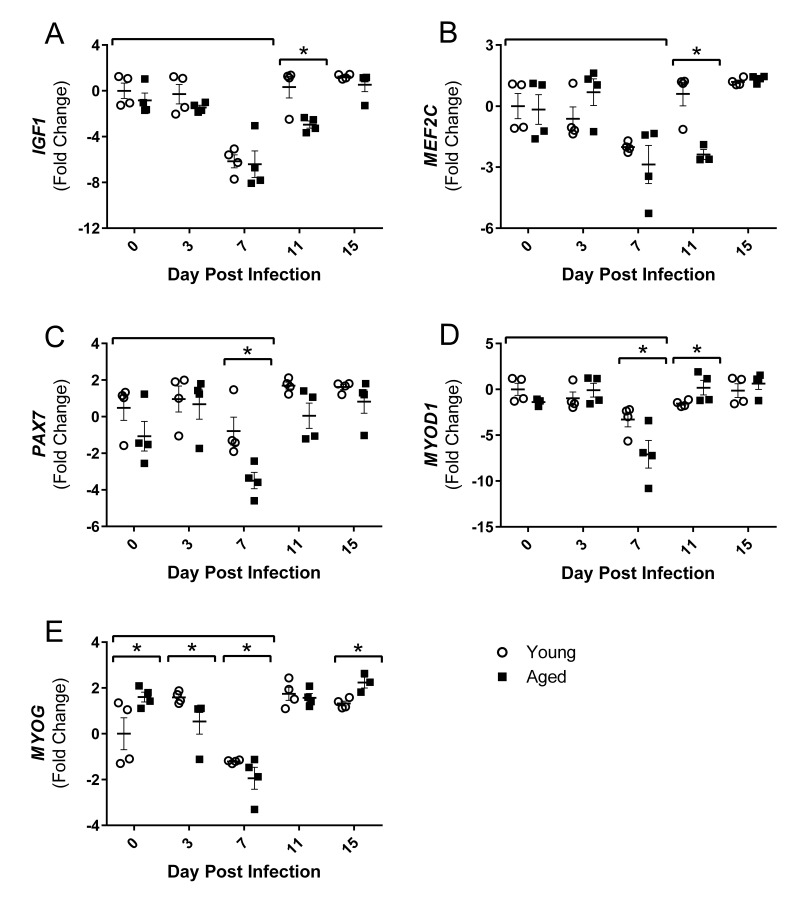
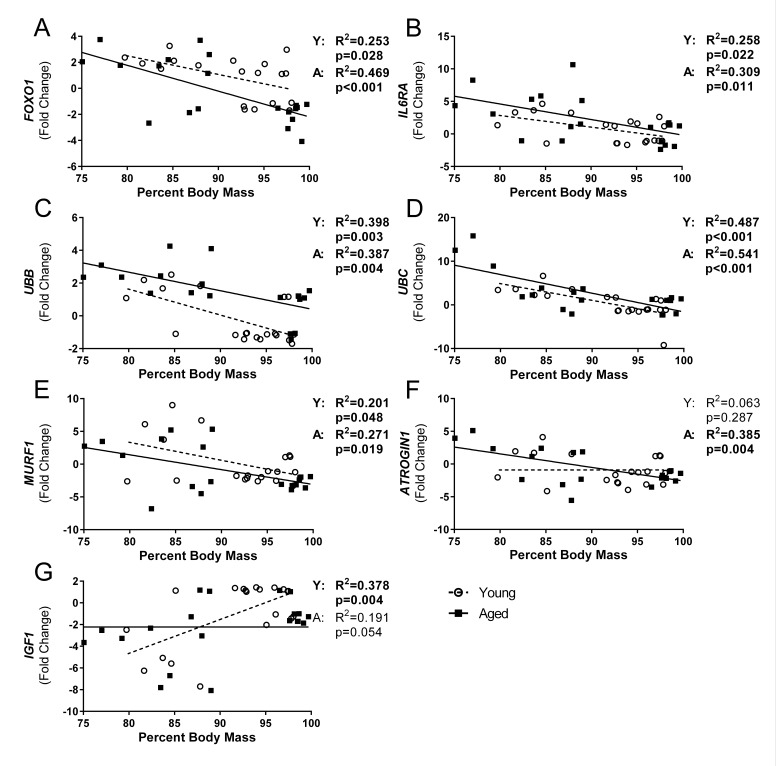
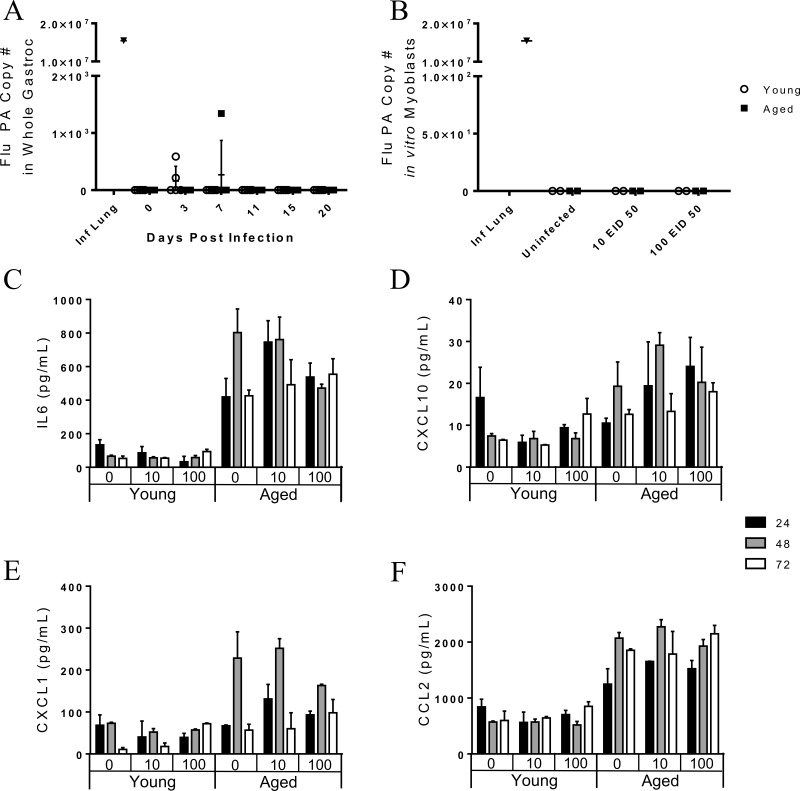
Although the influenza virus only infects the respiratory system, myalgias are commonly experienced during infection. In addition to a greater risk of hospitalization and death, older adults are more likely to develop disability following influenza infection; however, this relationship is understudied. We hypothesized that upon challenge with influenza, aging would be associated with functional impairments, as well as upregulation of skeletal muscle inflammatory and atrophy genes. Infected young and aged mice demonstrated decreased mobility and altered gait kinetics. These declines were more prominent in hind limbs and in aged mice. Skeletal muscle expression of genes involved in inflammation, as well as muscle atrophy and proteolysis, increased during influenza infection with an elevated and prolonged peak in aged mice. Infection also decreased expression of positive regulators of muscle mass and myogenesis components to a greater degree in aged mice. Gene expression correlated to influenza-induced body mass loss, although evidence did not support direct muscle infection. Overall, influenza leads to mobility impairments with induction of inflammatory and muscle degradation genes and downregulation of positive regulators of muscle. These effects are augmented and prolonged with aging, providing a molecular link between influenza infection, decreased resilience and increased risk of disability in the elderly.
Keywords: aging; disability; influenza; muscle atrophy; resilience.
Conflict of interest statement
The authors have no conflicting financial interests.
Figures
References
-
- Thompson WW, Shay DK, Weintraub E, Brammer L, Cox N, Anderson LJ, Fukuda K. Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA. 2003;289:179–186. - PubMed
-
- Heron M. Deaths: leading causes for 2010. Natl Vital Stat Rep. 2013;62:1–96. - PubMed
-
- Falsey AR, Hennessey PA, Formica MA, Cox C, Walsh EE. Respiratory syncytial virus infection in elderly and high-risk adults. The New England journal of medicine. 2005;352:1749–1759. - PubMed
-
- Ferrucci L, Guralnik JM, Pahor M, Corti MC, Havlik RJ. Hospital diagnoses, Medicare charges, and nursing home admissions in the year when older persons become severely disabled. JAMA. 1997;277:728–734. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
