

Low Concentrations of Nitric Oxide Modulate Streptococcus pneumoniae Biofilm Metabolism and Antibiotic Tolerance
- PMID: 26856845
- PMCID: PMC4808185
- DOI: 10.1128/AAC.02432-15
Low Concentrations of Nitric Oxide Modulate Streptococcus pneumoniae Biofilm Metabolism and Antibiotic Tolerance
Abstract
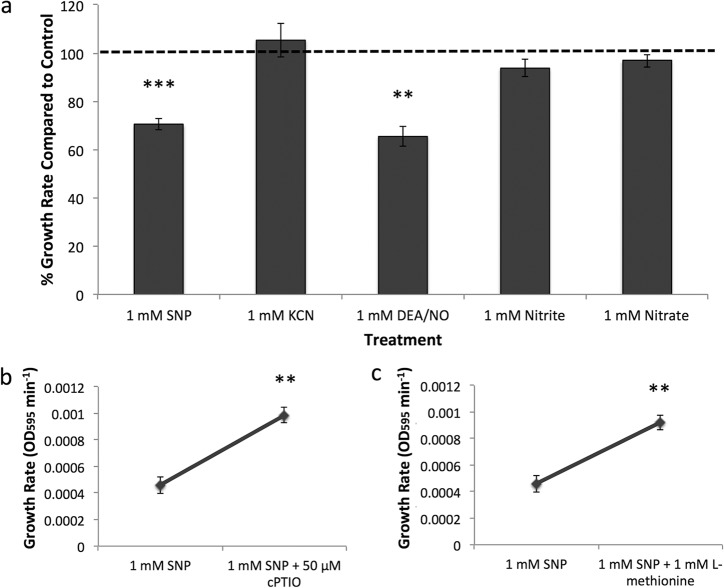
Streptococcus pneumoniaeis one of the key pathogens responsible for otitis media (OM), the most common infection in children and the largest cause of childhood antibiotic prescription. Novel therapeutic strategies that reduce the overall antibiotic consumption due to OM are required because, although widespread pneumococcal conjugate immunization has controlled invasive pneumococcal disease, overall OM incidence has not decreased. Biofilm formation represents an important phenotype contributing to the antibiotic tolerance and persistence ofS. pneumoniaein chronic or recurrent OM. We investigated the treatment of pneumococcal biofilms with nitric oxide (NO), an endogenous signaling molecule and therapeutic agent that has been demonstrated to trigger biofilm dispersal in other bacterial species. We hypothesized that addition of low concentrations of NO to pneumococcal biofilms would improve antibiotic efficacy and that higher concentrations exert direct antibacterial effects. Unlike in many other bacterial species, low concentrations of NO did not result inS. pneumoniaebiofilm dispersal. Instead, treatment of bothin vitrobiofilms andex vivoadenoid tissue samples (a reservoir forS. pneumoniaebiofilms) with low concentrations of NO enhanced pneumococcal killing when combined with amoxicillin-clavulanic acid, an antibiotic commonly used to treat chronic OM. Quantitative proteomic analysis using iTRAQ (isobaric tag for relative and absolute quantitation) identified 13 proteins that were differentially expressed following low-concentration NO treatment, 85% of which function in metabolism or translation. Treatment with low-concentration NO, therefore, appears to modulate pneumococcal metabolism and may represent a novel therapeutic approach to reduce antibiotic tolerance in pneumococcal biofilms.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures








References
-
- Vergison A, Dagan R, Arguedas A, Bonhoeffer J, Cohen R, Dhooge I, Hoberman A, Liese J, Marchisio P, Palmu AA, Ray GT, Sanders EAM, Simoes EA, Uhari M, van Eldere J, Pelton SI. 2010. Otitis media and its consequences: beyond the earache. Lancet Infect Dis 10:195–203. doi:10.1016/S1473-3099(10)70012-8. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
