

A Signaling Cascade from miR444 to RDR1 in Rice Antiviral RNA Silencing Pathway
- PMID: 26858364
- PMCID: PMC4825140
- DOI: 10.1104/pp.15.01283
A Signaling Cascade from miR444 to RDR1 in Rice Antiviral RNA Silencing Pathway
Abstract
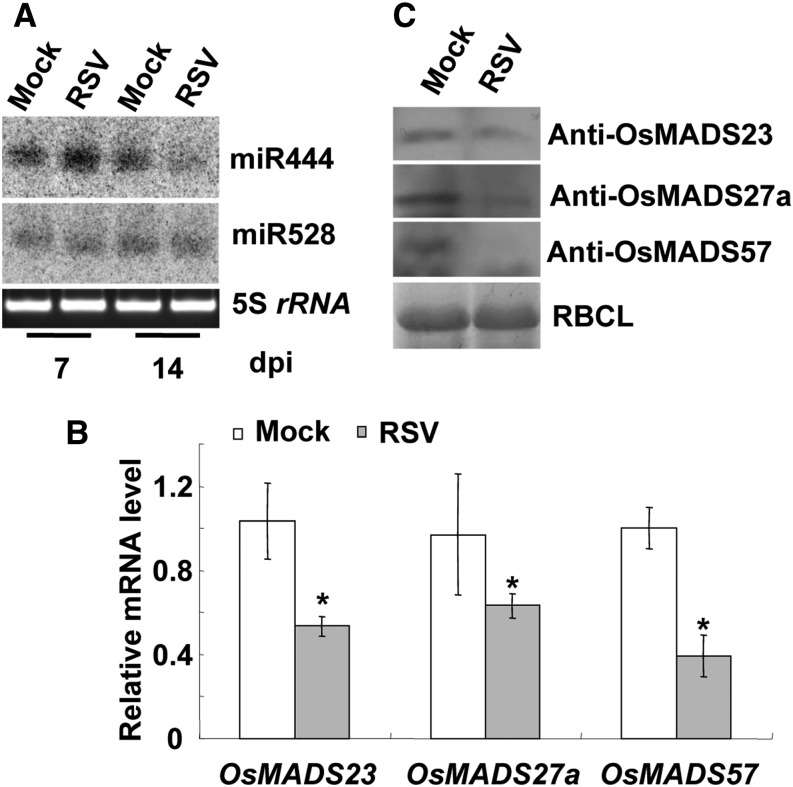
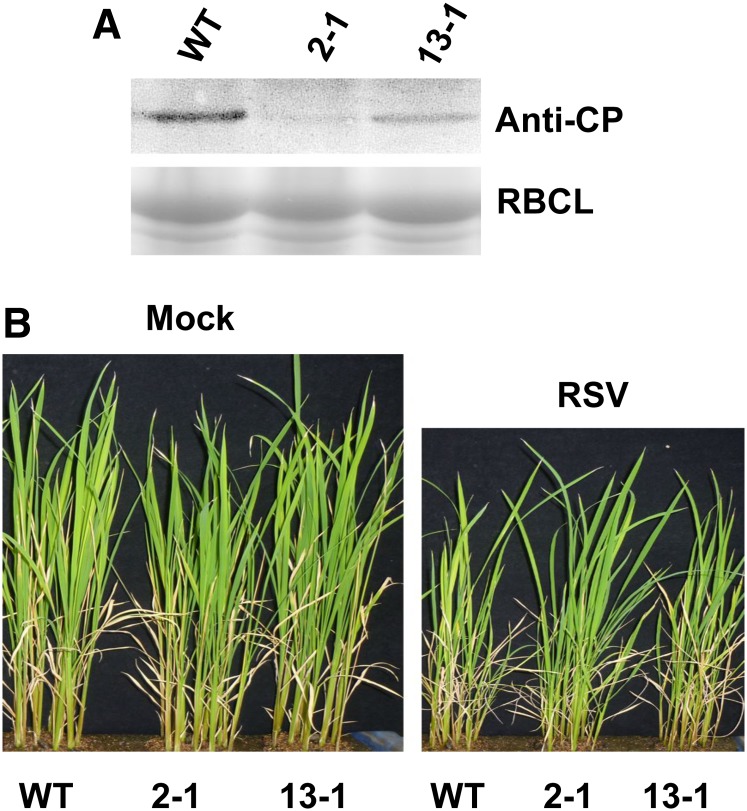
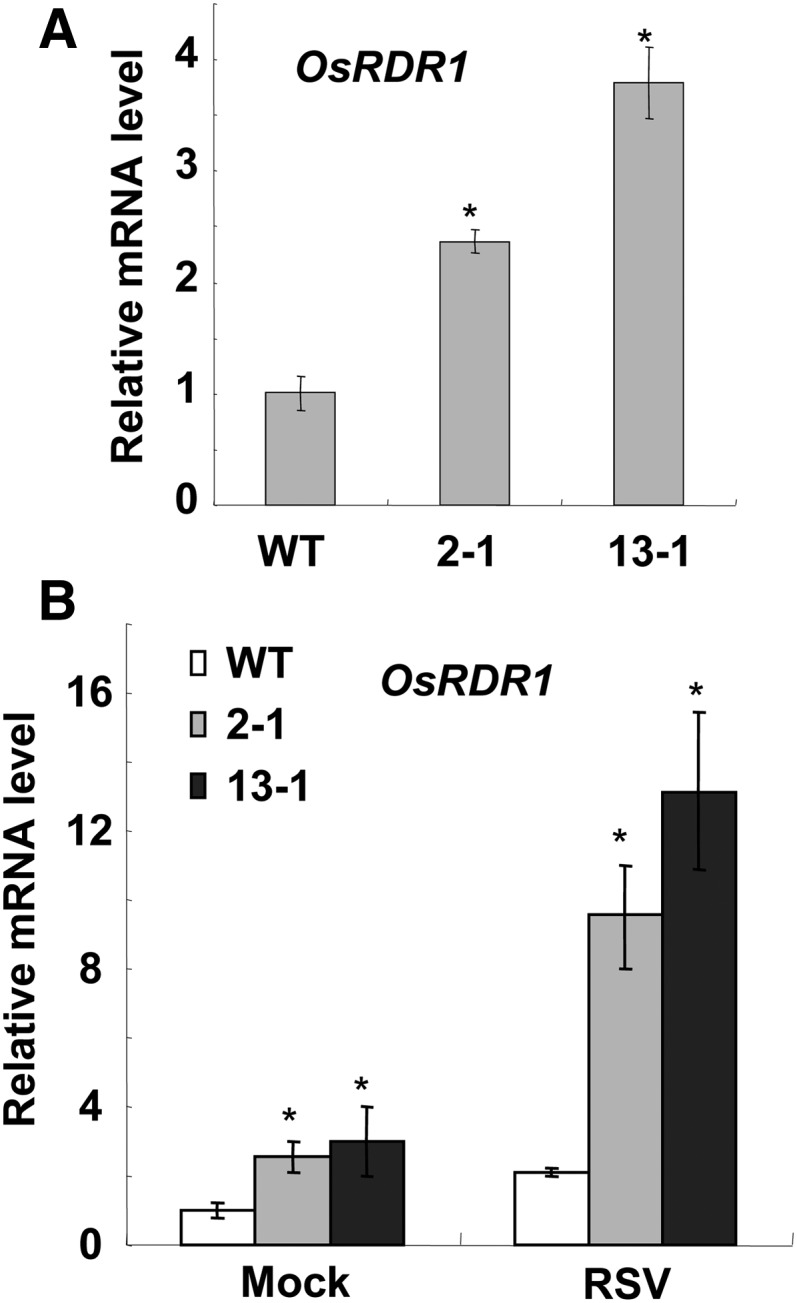
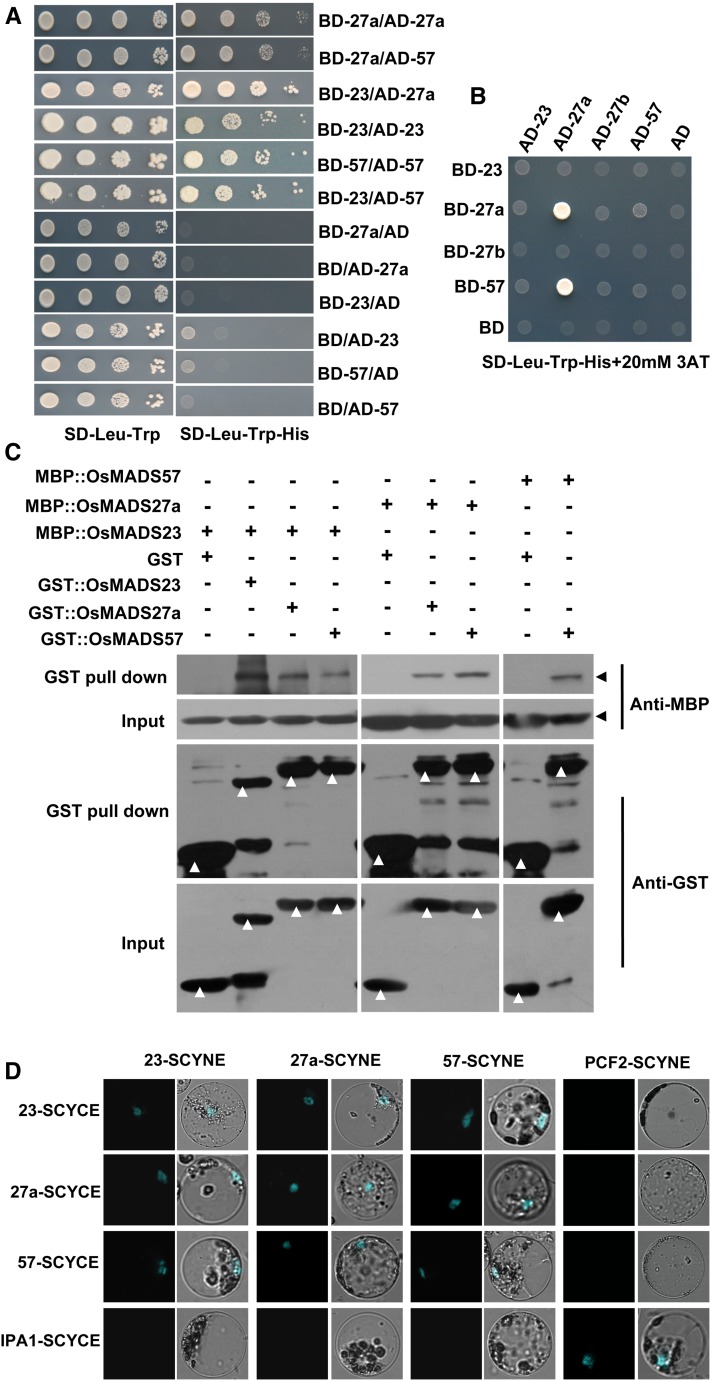
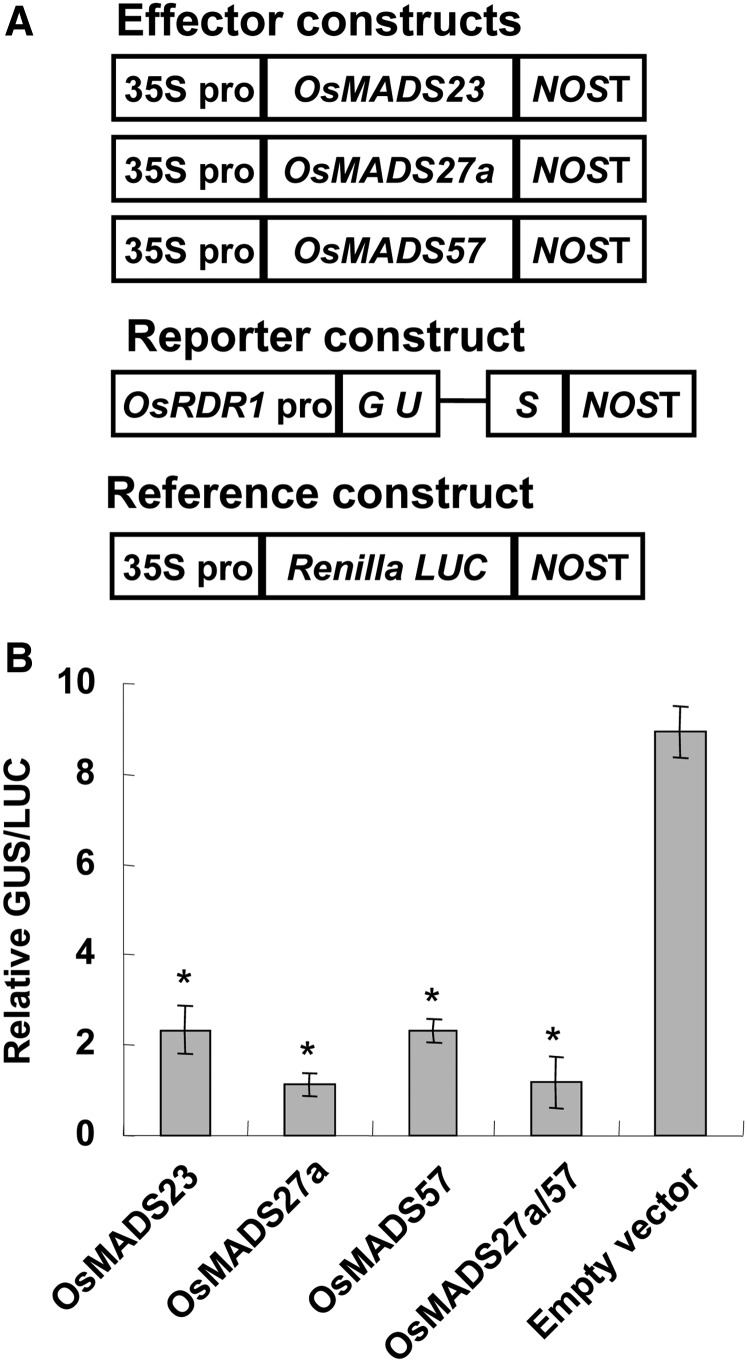
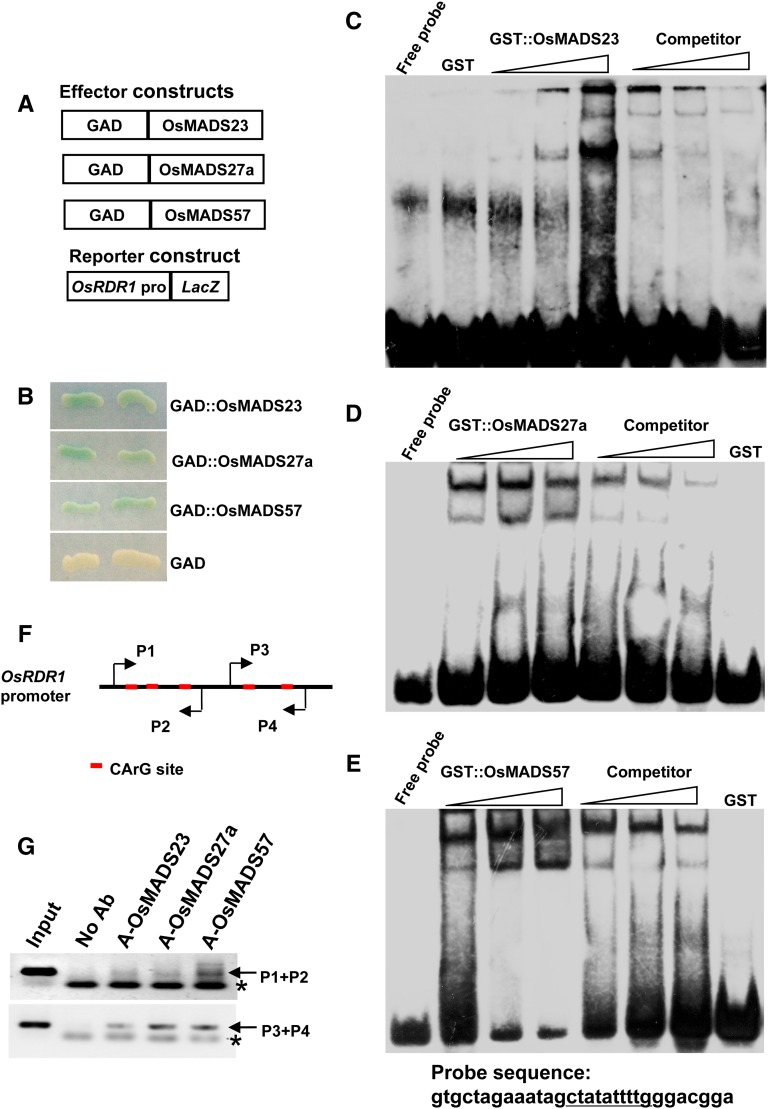
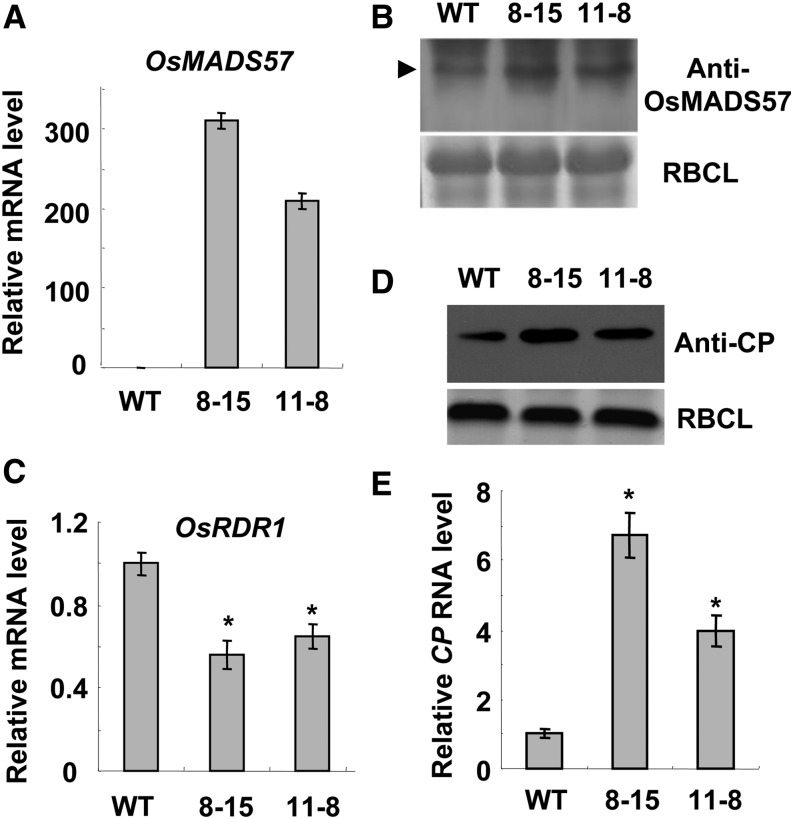
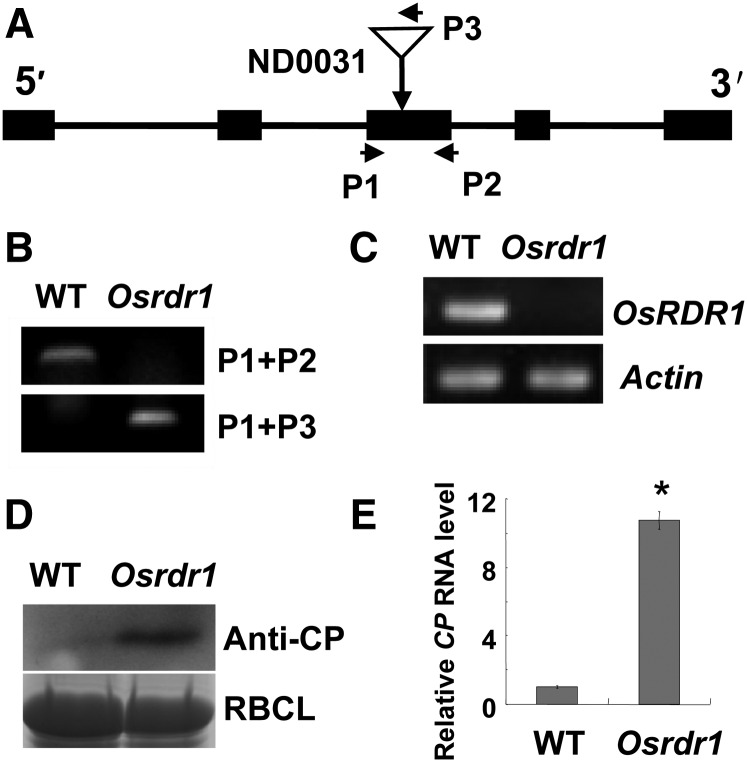
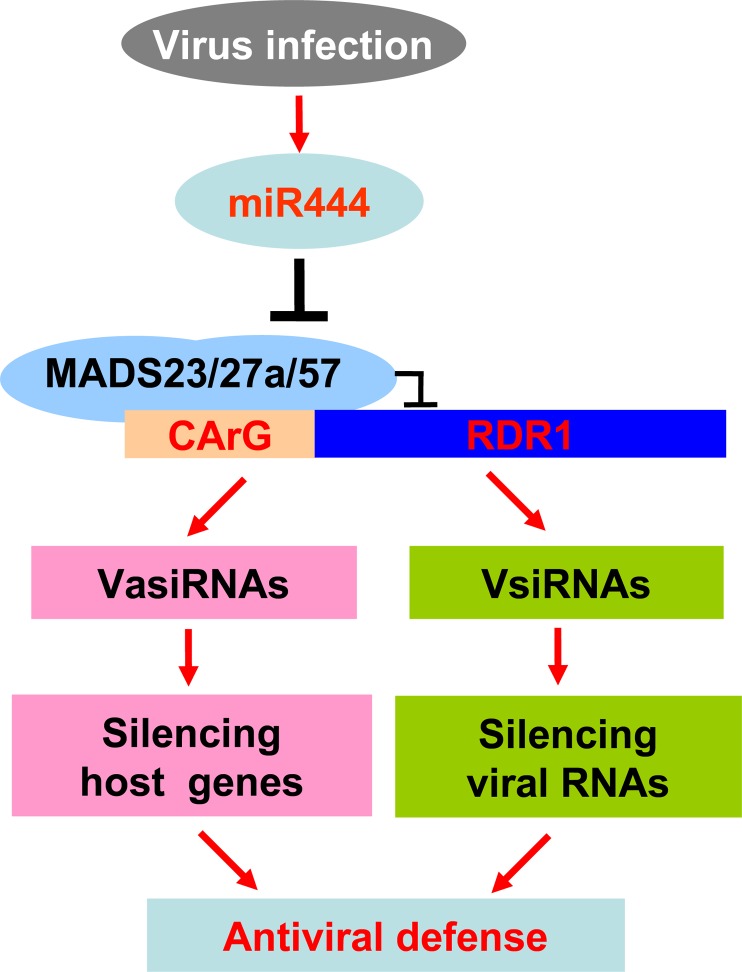
Plant RNA-DEPENDENT RNA POLYMERASE1 (RDR1) is a key component of the antiviral RNA-silencing pathway, contributing to the biogenesis of virus-derived small interfering RNAs. This enzyme also is responsible for producing virus-activated endogenous small interfering RNAs to stimulate the broad-spectrum antiviral activity through silencing host genes. The expression of RDR1 orthologs in various plants is usually induced by virus infection. However, the molecular mechanisms of activation of RDR1 expression in response to virus infection remain unknown. Here, we show that a monocot-specific microRNA, miR444, is a key factor in relaying the antiviral signaling from virus infection to OsRDR1 expression. The expression of miR444 is enhanced by infection with Rice stripe virus (RSV), and overexpression of miR444 improves rice (Oryza sativa) resistance against RSV infection accompanied by the up-regulation of OsRDR1 expression. We further show that three miR444 targets, the MIKC(C)-type MADS box proteins OsMADS23, OsMADS27a, and OsMADS57, form homodimers and heterodimers between them to repress the expression of OsRDR1 by directly binding to the CArG motifs of its promoter. Consequently, an increased level of miR444 diminishes the repressive roles of OsMADS23, OsMADS27a, and OsMADS57 on OsRDR1 transcription, thus activating the OsRDR1-dependent antiviral RNA-silencing pathway. We also show that overexpression of miR444-resistant OsMADS57 reduced OsRDR1 expression and rice resistance against RSV infection, and knockout of OsRDR1 reduced rice resistance against RSV infection. In conclusion, our results reveal a molecular cascade in the rice antiviral pathway in which miR444 and its MADS box targets directly control OsRDR1 transcription.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures
Similar articles
-
Mutation of the RDR1 gene caused genome-wide changes in gene expression, regional variation in small RNA clusters and localized alteration in DNA methylation in rice.BMC Plant Biol. 2014 Jun 30;14:177. doi: 10.1186/1471-2229-14-177. BMC Plant Biol. 2014. PMID: 24980094 Free PMC article.
-
Ferredoxin 1 is downregulated by the accumulation of abscisic acid in an ABI5-dependent manner to facilitate rice stripe virus infection in Nicotiana benthamiana and rice.Plant J. 2021 Aug;107(4):1183-1197. doi: 10.1111/tpj.15377. Epub 2021 Jul 10. Plant J. 2021. PMID: 34153146
-
The hypersensitive induced reaction 3 (HIR3) gene contributes to plant basal resistance via an EDS1 and salicylic acid-dependent pathway.Plant J. 2019 Jun;98(5):783-797. doi: 10.1111/tpj.14271. Epub 2019 Feb 28. Plant J. 2019. PMID: 30730076
-
Recent progress on gene silencing/suppression by virus-derived small interfering RNAs in rice viruses especially Rice grassy stunt virus.Microb Pathog. 2018 Dec;125:210-218. doi: 10.1016/j.micpath.2018.09.021. Epub 2018 Sep 19. Microb Pathog. 2018. PMID: 30243549 Review.
-
Antiviral roles of plant ARGONAUTES.Curr Opin Plant Biol. 2015 Oct;27:111-7. doi: 10.1016/j.pbi.2015.06.013. Epub 2015 Jul 17. Curr Opin Plant Biol. 2015. PMID: 26190744 Free PMC article. Review.
Cited by
-
A Review of Vector-Borne Rice Viruses.Viruses. 2022 Oct 14;14(10):2258. doi: 10.3390/v14102258. Viruses. 2022. PMID: 36298813 Free PMC article. Review.
-
Plant Noncoding RNAs: Hidden Players in Development and Stress Responses.Annu Rev Cell Dev Biol. 2019 Oct 6;35:407-431. doi: 10.1146/annurev-cellbio-100818-125218. Epub 2019 Aug 12. Annu Rev Cell Dev Biol. 2019. PMID: 31403819 Free PMC article. Review.
-
Evolution of Disease Defense Genes and Their Regulators in Plants.Int J Mol Sci. 2019 Jan 15;20(2):335. doi: 10.3390/ijms20020335. Int J Mol Sci. 2019. PMID: 30650550 Free PMC article. Review.
-
Characterization of Maize miRNAs in Response to Synergistic Infection of Maize Chlorotic Mottle Virus and Sugarcane Mosaic Virus.Int J Mol Sci. 2019 Jun 27;20(13):3146. doi: 10.3390/ijms20133146. Int J Mol Sci. 2019. PMID: 31252649 Free PMC article.
-
Traversing the Links between Heavy Metal Stress and Plant Signaling.Front Plant Sci. 2018 Feb 5;9:12. doi: 10.3389/fpls.2018.00012. eCollection 2018. Front Plant Sci. 2018. PMID: 29459874 Free PMC article. Review.
References
-
- Alamillo JM, Saénz P, García JA (2006) Salicylic acid-mediated and RNA-silencing defense mechanisms cooperate in the restriction of systemic spread of plum pox virus in tobacco. Plant J 48: 217–227 - PubMed
-
- Axtell MJ. (2013) Classification and comparison of small RNAs from plants. Annu Rev Plant Biol 64: 137–159 - PubMed
-
- Bologna NG, Voinnet O (2014) The diversity, biogenesis, and activities of endogenous silencing small RNAs in Arabidopsis. Annu Rev Plant Biol 65: 473–503 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
