

Transient Glyco-Engineering to Produce Recombinant IgA1 with Defined N- and O-Glycans in Plants
- PMID: 26858738
- PMCID: PMC4731523
- DOI: 10.3389/fpls.2016.00018
Transient Glyco-Engineering to Produce Recombinant IgA1 with Defined N- and O-Glycans in Plants
Abstract
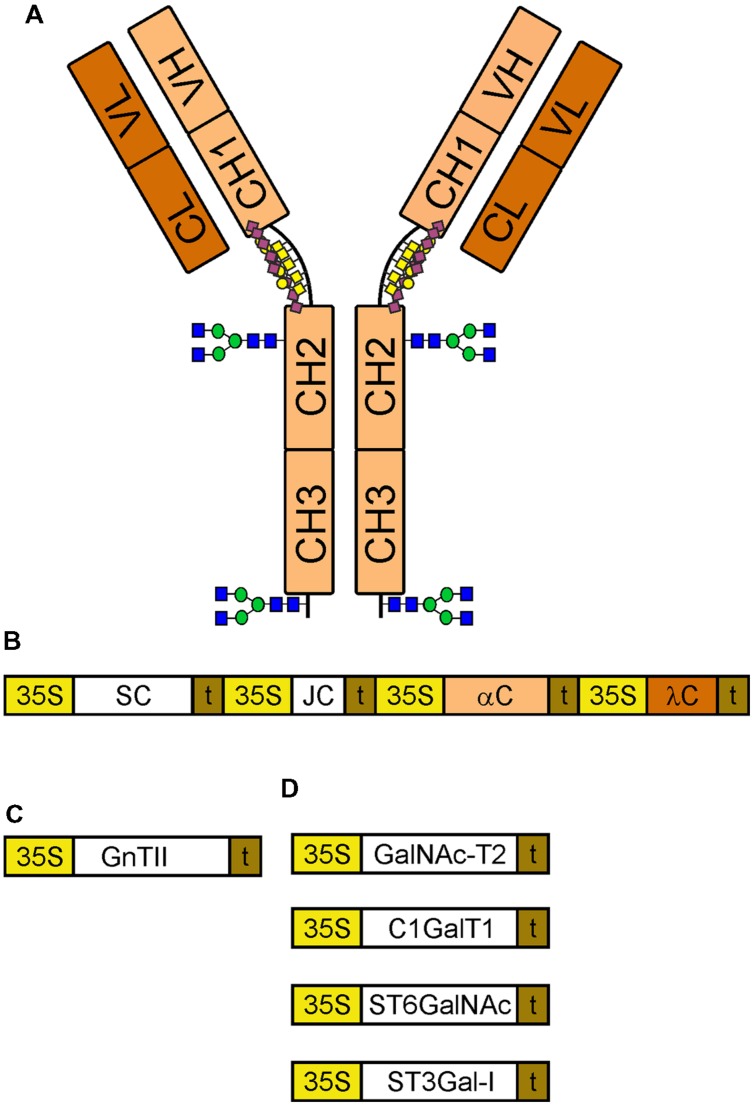
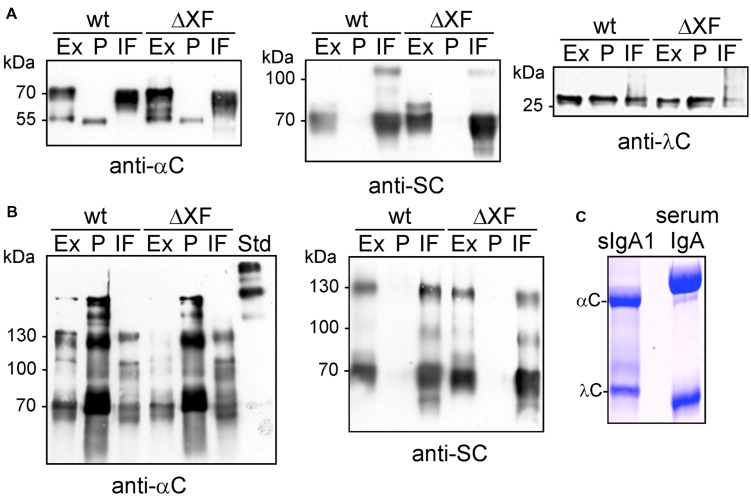
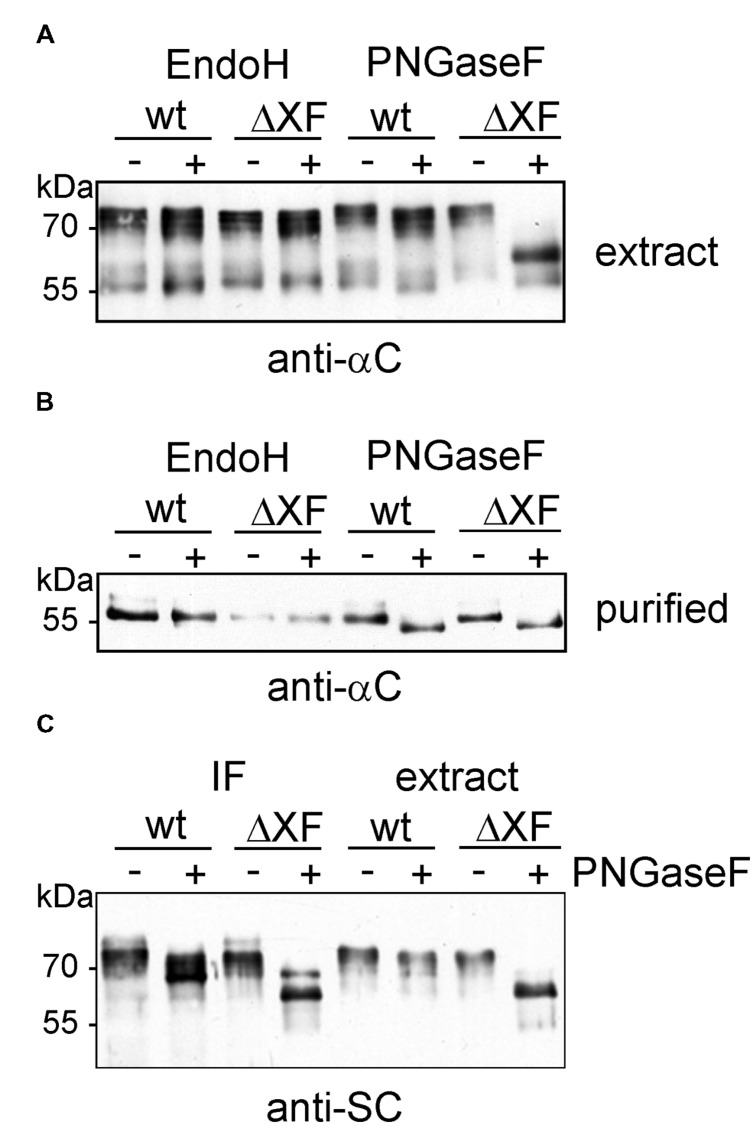
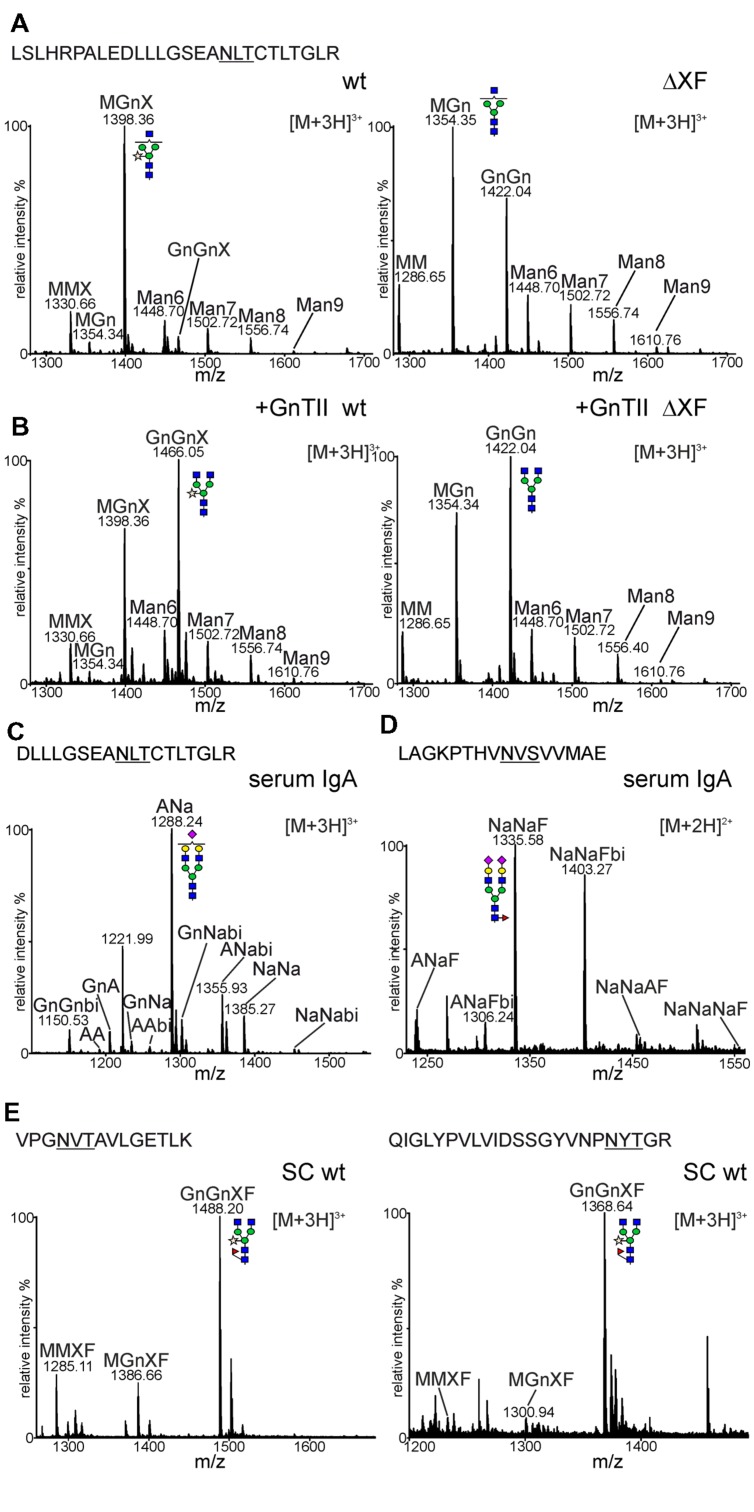
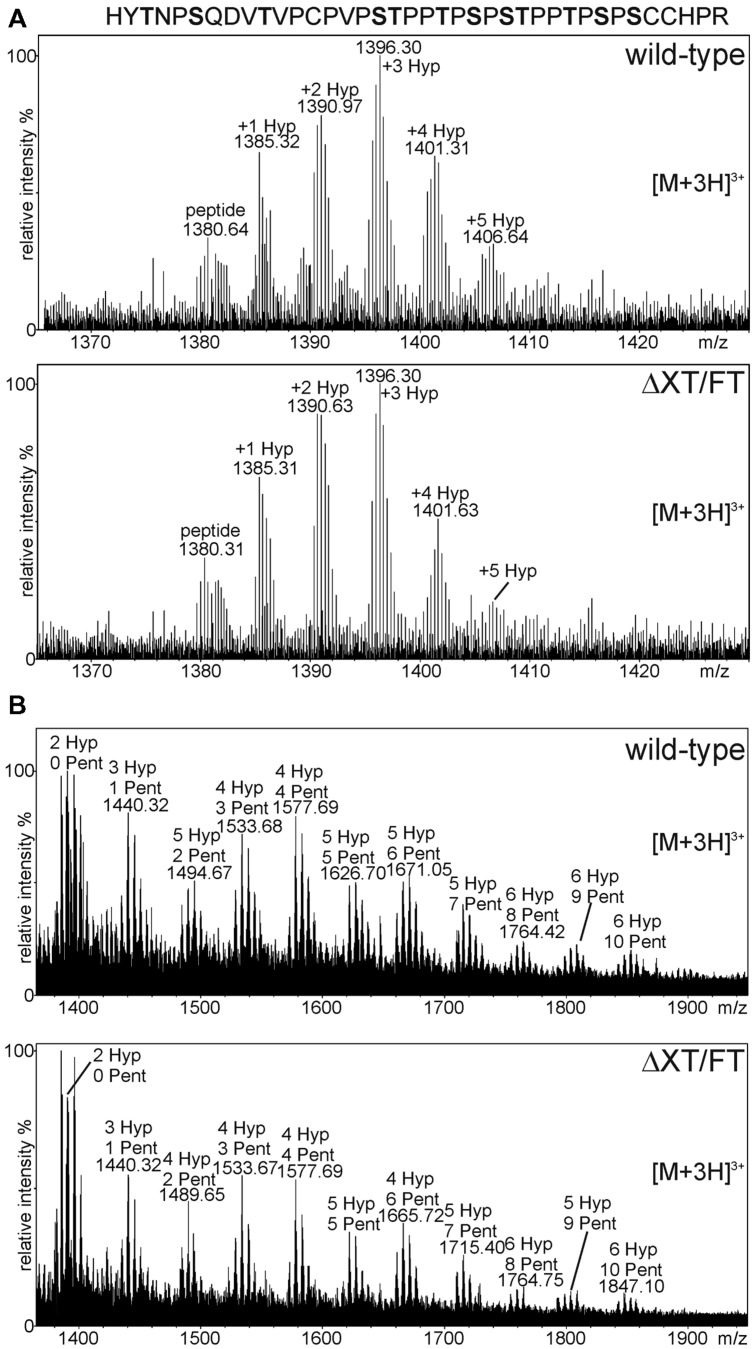
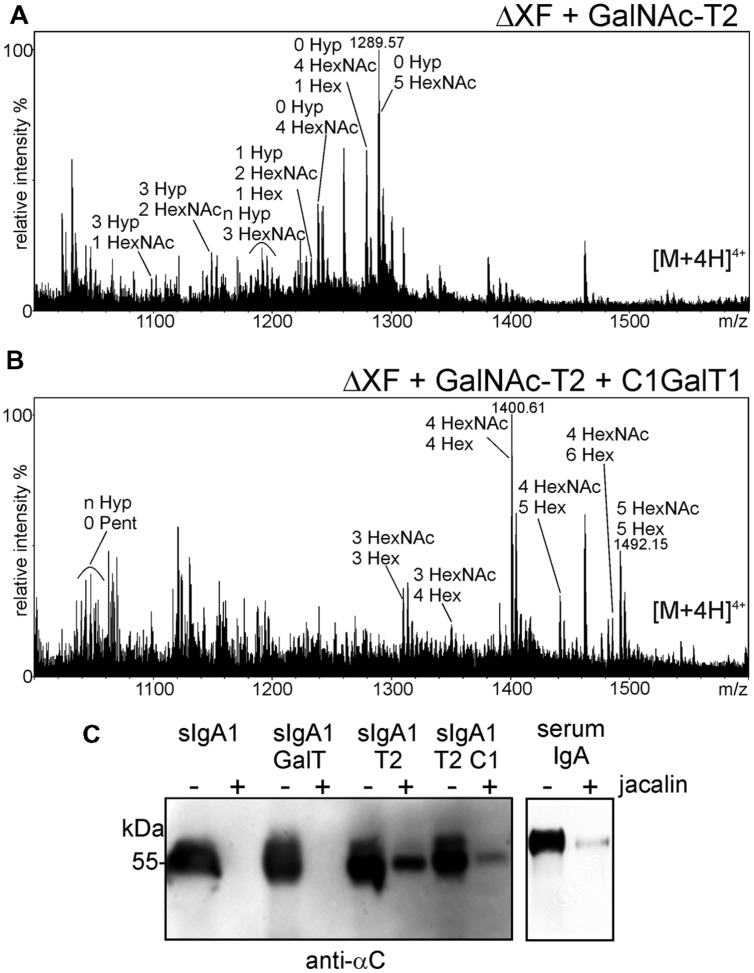
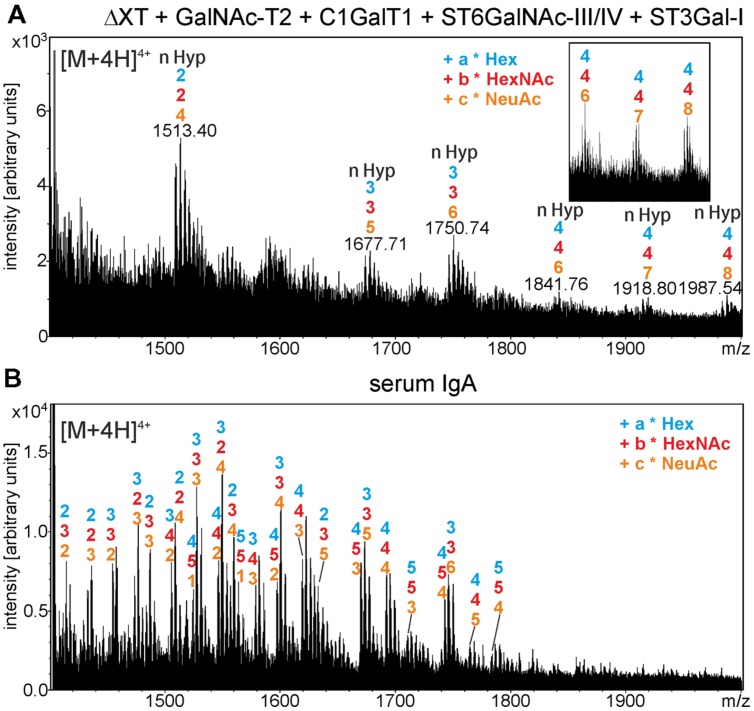
The production of therapeutic antibodies to combat pathogens and treat diseases, such as cancer is of great interest for the biotechnology industry. The recent development of plant-based expression systems has demonstrated that plants are well-suited for the production of recombinant monoclonal antibodies with defined glycosylation. Compared to immunoglobulin G (IgG), less effort has been undertaken to express immunoglobulin A (IgA), which is the most prevalent antibody class at mucosal sites and a promising candidate for novel recombinant biopharmaceuticals with enhanced anti-tumor activity. Here, we transiently expressed recombinant human IgA1 against the VP8* rotavirus antigen in glyco-engineered ΔXT/FT Nicotiana benthamiana plants. Mass spectrometric analysis of IgA1 glycopeptides revealed the presence of complex biantennary N-glycans with terminal N-acetylglucosamine present on the N-glycosylation site of the CH2 domain in the IgA1 alpha chain. Analysis of the peptide carrying nine potential O-glycosylation sites in the IgA1 alpha chain hinge region showed the presence of plant-specific modifications including hydroxyproline formation and the attachment of pentoses. By co-expression of enzymes required for initiation and elongation of human O-glycosylation it was possible to generate disialylated mucin-type core 1 O-glycans on plant-produced IgA1. Our data demonstrate that ΔXT/FT N. benthamiana plants can be engineered toward the production of recombinant IgA1 with defined human-type N- and O-linked glycans.
Keywords: N-glycosylation; O-glycosylation; antibody; glyco-engineering; monomeric IgA; plant-made pharmaceuticals; protein glycosylation; recombinant glycoprotein.
Figures
Similar articles
-
Exploring Site-Specific N-Glycosylation of HEK293 and Plant-Produced Human IgA Isotypes.J Proteome Res. 2017 Jul 7;16(7):2560-2570. doi: 10.1021/acs.jproteome.7b00121. Epub 2017 May 26. J Proteome Res. 2017. PMID: 28516782 Free PMC article.
-
Glyco-engineering for the production of recombinant IgA1 with distinct mucin-type O-glycans in plants.Bioengineered. 2016 Nov;7(6):484-489. doi: 10.1080/21655979.2016.1201251. Epub 2016 Jun 22. Bioengineered. 2016. PMID: 27333379 Free PMC article.
-
Engineering of sialylated mucin-type O-glycosylation in plants.J Biol Chem. 2012 Oct 19;287(43):36518-26. doi: 10.1074/jbc.M112.402685. Epub 2012 Sep 4. J Biol Chem. 2012. PMID: 22948156 Free PMC article.
-
The pathogenic role of IgA1 O-linked glycosylation in the pathogenesis of IgA nephropathy.Nephrology (Carlton). 2007 Jun;12(3):275-84. doi: 10.1111/j.1440-1797.2007.00797.x. Nephrology (Carlton). 2007. PMID: 17498123 Review.
-
IgA nephropathy and Henoch-Schoenlein purpura nephritis: aberrant glycosylation of IgA1, formation of IgA1-containing immune complexes, and activation of mesangial cells.Contrib Nephrol. 2007;157:134-8. doi: 10.1159/000102455. Contrib Nephrol. 2007. PMID: 17495451 Review.
Cited by
-
A Roadmap for the Molecular Farming of Viral Glycoprotein Vaccines: Engineering Glycosylation and Glycosylation-Directed Folding.Front Plant Sci. 2020 Dec 3;11:609207. doi: 10.3389/fpls.2020.609207. eCollection 2020. Front Plant Sci. 2020. PMID: 33343609 Free PMC article. Review.
-
Plant-derived secretory component gives protease-resistance to Shiga toxin 1-specific dimeric IgA.Plant Mol Biol. 2021 Jun;106(3):297-308. doi: 10.1007/s11103-021-01151-x. Epub 2021 Apr 19. Plant Mol Biol. 2021. PMID: 33871797 No abstract available.
-
Improved CRISPR/Cas9 gene editing by fluorescence activated cell sorting of green fluorescence protein tagged protoplasts.BMC Biotechnol. 2019 Jun 17;19(1):36. doi: 10.1186/s12896-019-0530-x. BMC Biotechnol. 2019. PMID: 31208390 Free PMC article.
-
Exploring Site-Specific N-Glycosylation of HEK293 and Plant-Produced Human IgA Isotypes.J Proteome Res. 2017 Jul 7;16(7):2560-2570. doi: 10.1021/acs.jproteome.7b00121. Epub 2017 May 26. J Proteome Res. 2017. PMID: 28516782 Free PMC article.
-
Investigation of a monoclonal antibody against enterotoxigenic Escherichia coli, expressed as secretory IgA1 and IgA2 in plants.Gut Microbes. 2021 Jan-Dec;13(1):1-14. doi: 10.1080/19490976.2020.1859813. Gut Microbes. 2021. PMID: 33439092 Free PMC article.
References
-
- Atkin J. D., Pleass R. J., Owens R. J., Woof J. M. (1996). Mutagenesis of the human IgA1 heavy chain tailpiece that prevents dimer assembly. J. Immunol. 157 156–159. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous